File: <culicidaekey.htm> <Medical Index> <General Index> Site Description Glossary <Navigate to Home>
|
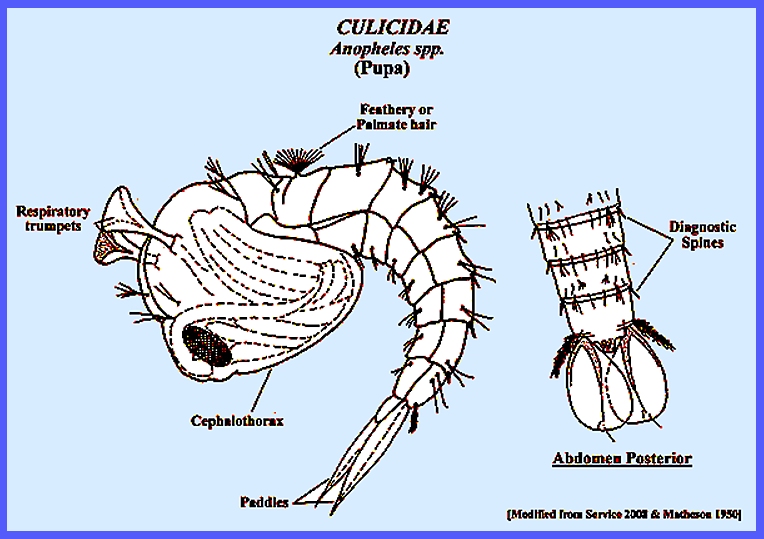
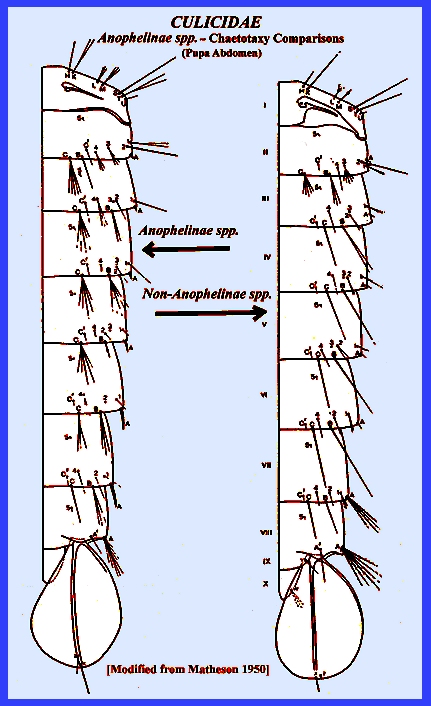
Insecta: Diptera CULICIDAE KEY (Mosquitoes) (Subfamilies, Tribes, Genera,
Subgenera) (Contact) Please CLICK on picture and
underlined links to view or to navigate within the key: To Search for
Subject Matter use Ctrl/F There are over 3,425
species of mosquitoes in 43 or more genera described as of 2016. There continues to be disagreement among
specialists on the number of genera.
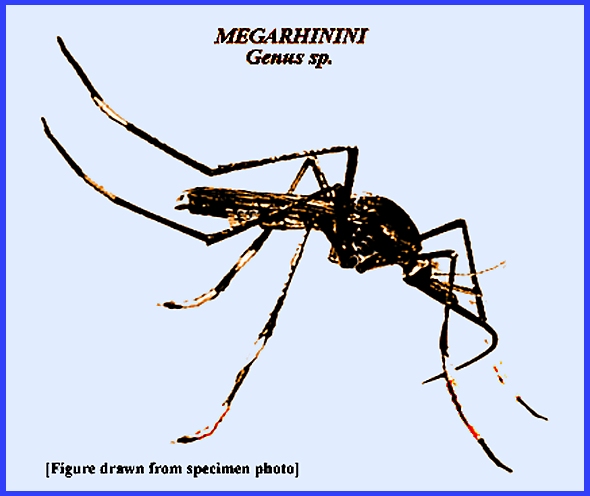
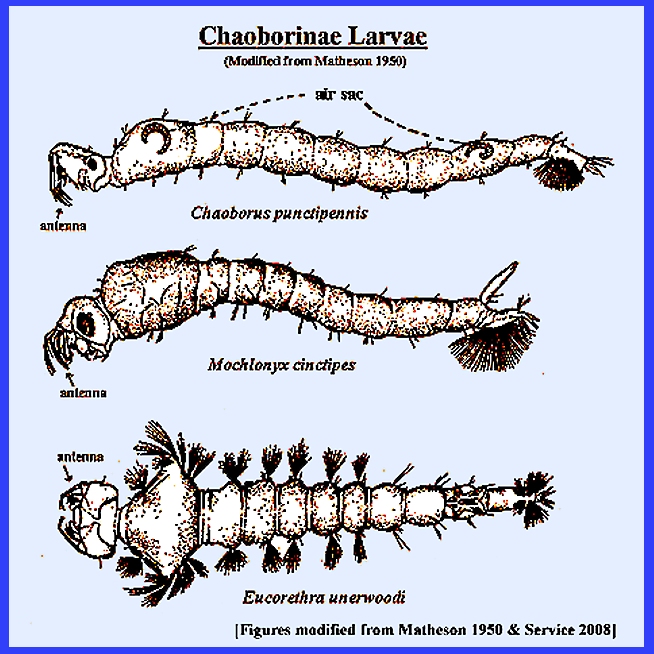
Matheson (1950) lists two subfamilies: Chaoborinae and Culicinae and then further separates the group
into four tribes: Anophilini, Culicini, Magarhinini and Sabethini. Service (2008) has three subfamilies: Anophelini, Culicinae. and Toxorhynchitinae. There is only one genus in
the Toxorhynchitinae (Toxorhynchites)
the members of which do not bite and thus are of no medical importance even
though their larvae are predators of other mosquitoes and may be considered
beneficial. The Culicidae are
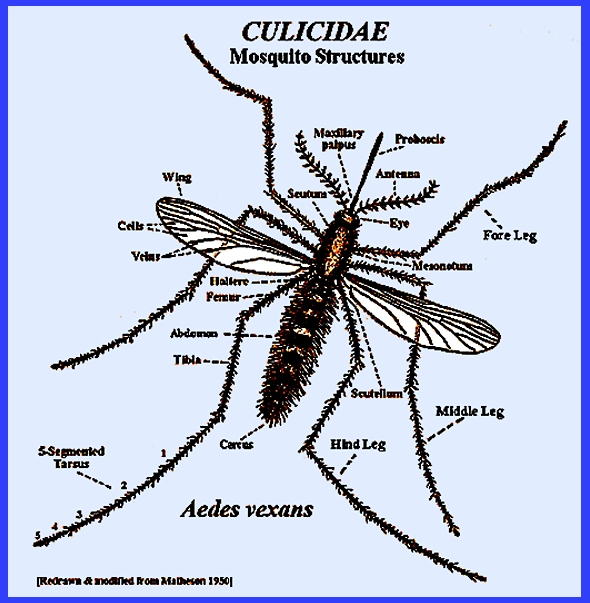
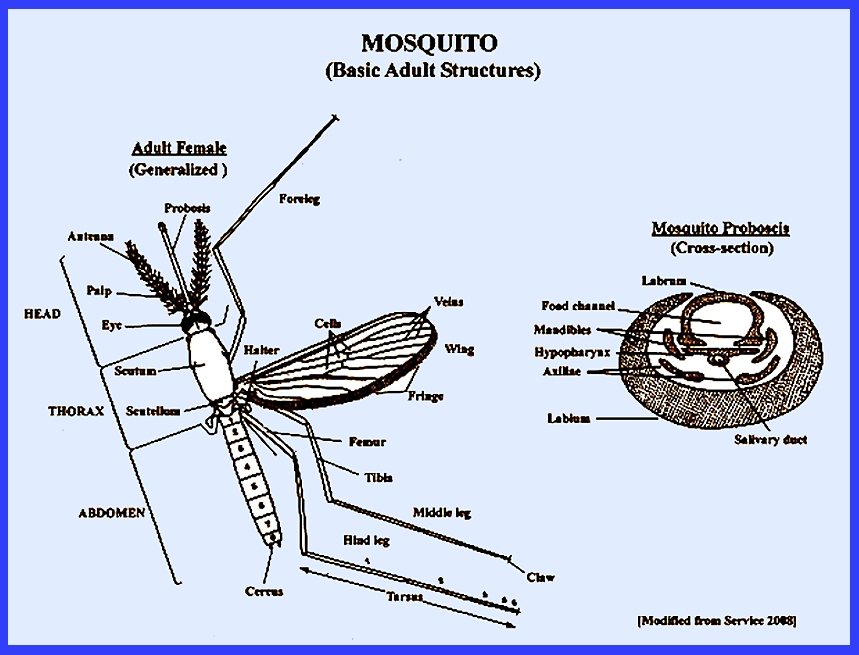
distinguished by having only one pair of functional fore wings, with the hind
wings modified as halteres. There is
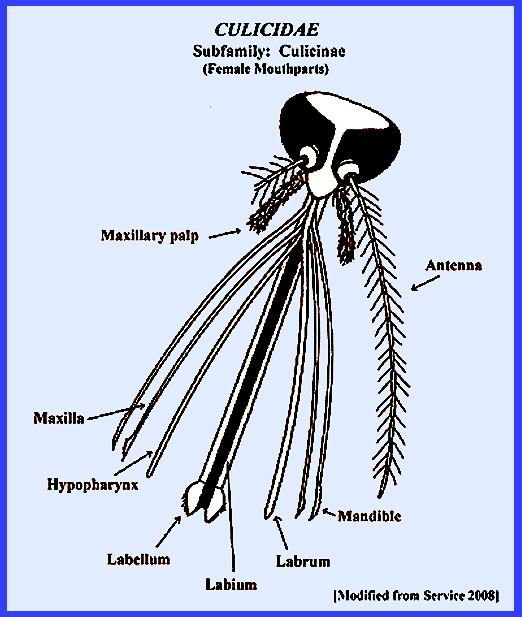
a forward-projecting proboscis, many scales on the thorax, legs, abdomen and
wing veins and a fringe of scales along the posterior margin of the
wings. Most species are small and
slender. The eyes are
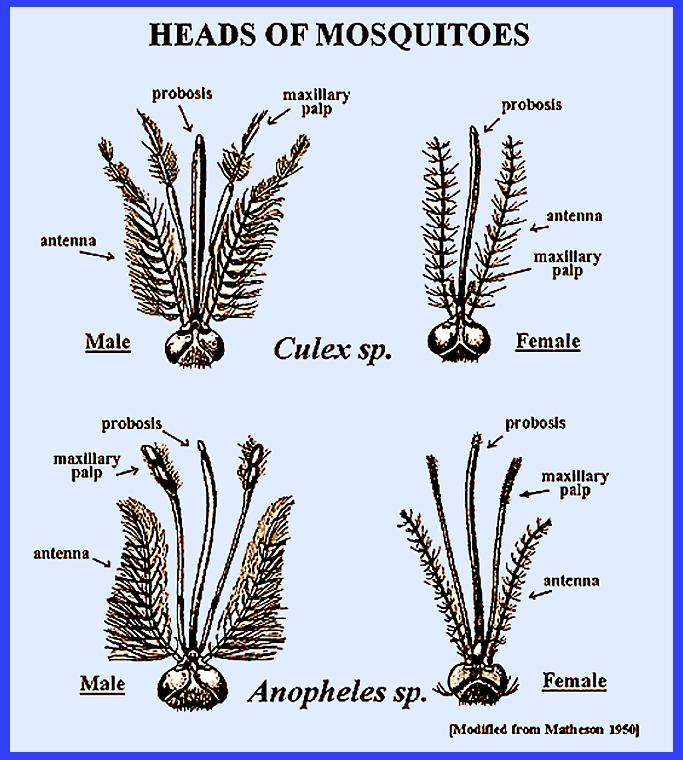
kidney-shaped. The antennae are
segmented and have short hairs in females and long hairs in males. The following keys use
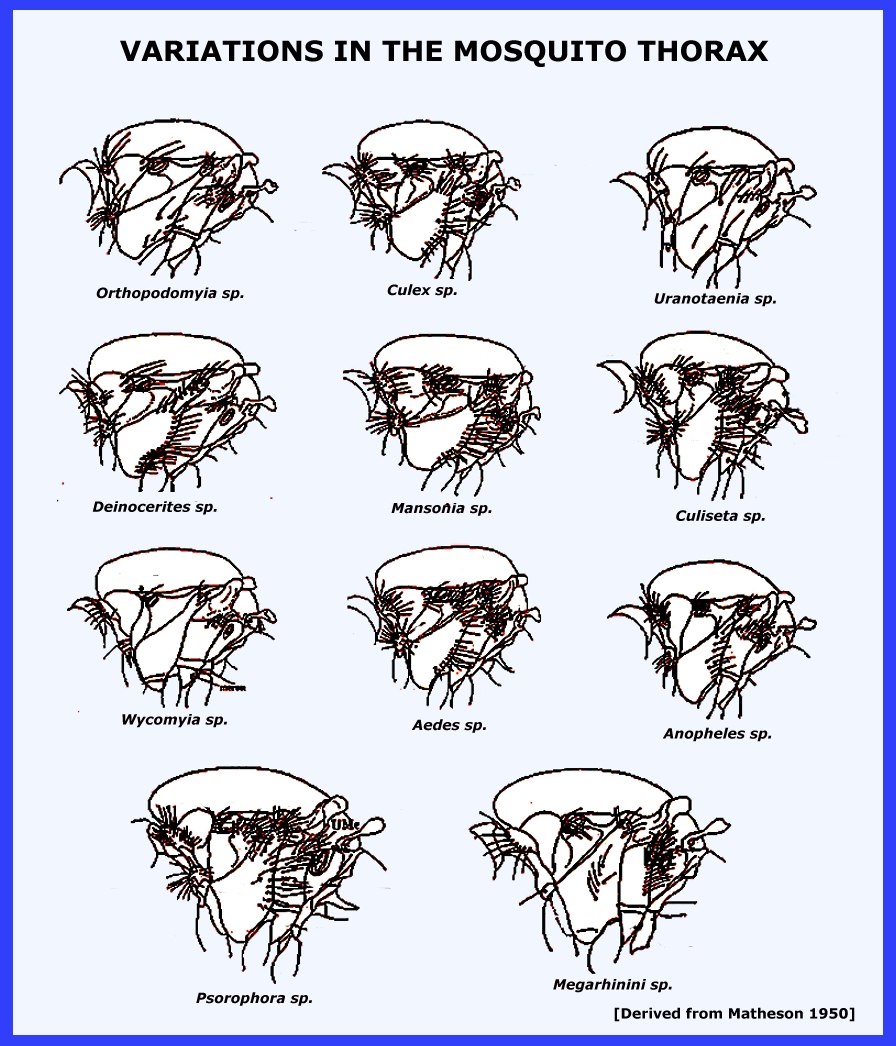
the Matheson (1950) system to separate subfamilies, tribes, genera and
subgenera in order to facilitate the identification of species in this very
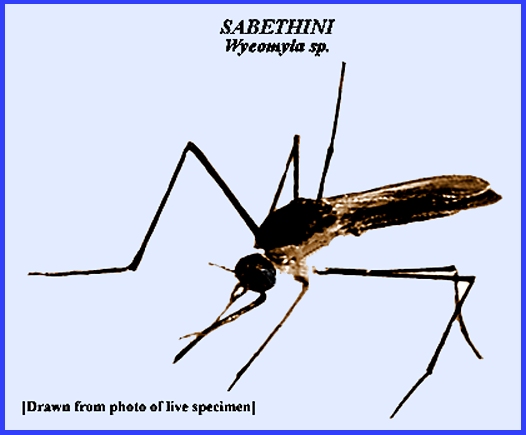
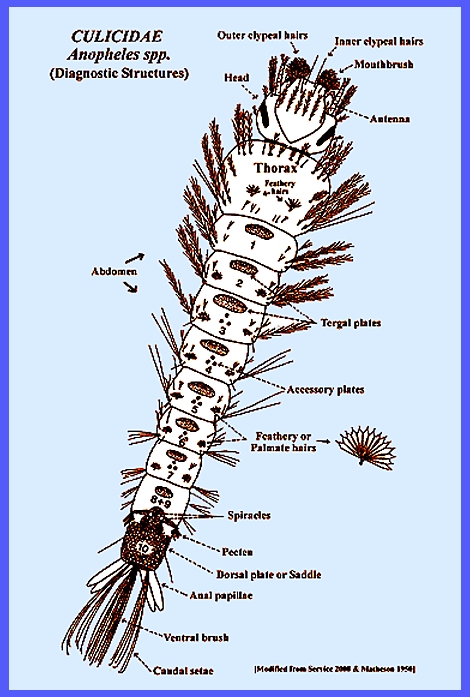
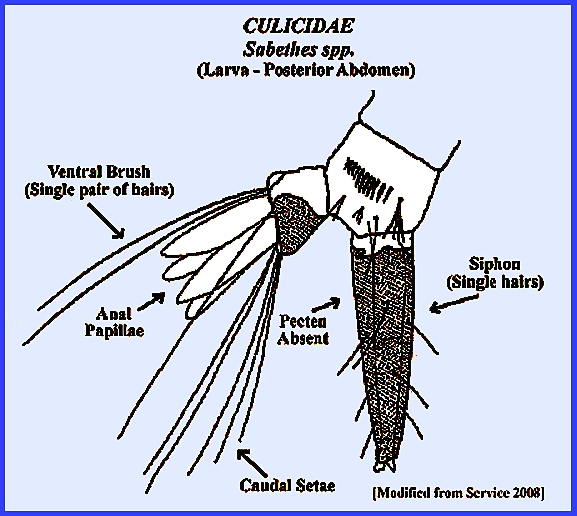
large and diverse family Culicidae. Navigate directly to: Subfamilies, Tribes, Anophilini Genera, Culicini Genera-adults, Culicini Genera-larvae, North American Anophelines-adults, North American Anophelines-larvae. Because the Matheson
arrangement includes taxa that bear a close resemblance to Culicidae it is
well suited to the nonspecialist for it distinguishes the medically important
groups from the more benign. = =
= = = = = = = = = = = = = = = = = = = =
_ _ _ _ _ _
_ _ _ _ _ _ _ _ _ _ _ _ _ _ Key References: <medvet.ref.htm> <Hexapoda> Bock, G. R. & G.
Cardew. 1996. Olfaction in Mosquito-Host
Interactions. Chichester: Wiley
Publ., England Clark, G. G.
1994. Prevention of tropical
diseases: status of new and emerging vector control strategies. Proc. Symp. Vector Control, Amer. J. Trop.
Med. & Hyg. 50(6): 1-159. Clements, A. N. 1992.
The Biology of Mosquitoes. Vol. 1:
Development, Nutrition & Reproduction, Chapman & Hall, London. Curtis, C. F. 1989.
Appropriate Technology in Vector Control. CRC Press, Boca Raton, Florida. Foster, W. A. & E.
D. Walker. 2002. Mosquitoes (Culicidae). IN: Med. &
Veterinary Ent.. Acad. Press,
Amsterdam. pp. 203-62. Matheson, R. 1950. Medical Entomology. Comstock Publ. Co, Inc. 610 p. Service, M. 2008.
Medical Entomology For Students.
Cambridge Univ. Press. 289 p Legner, E. F. 1995. Biological control of Diptera of medical and veterinary
importance. J. Vector Ecology 20(1):
59-120. Legner, E. F.. 2000. Biological control of aquatic Diptera. p. 847-870. Contributions to a Manual of Palaearctic Diptera, Vol. 1, Sci.
Herald, Budapest. 978 p. Pates, H. & C. Curtis. 2005. Mosquito behavior and vector control. Ann. Rev. Ent. 50:
53-70. Spielman, A. &
M. d'Antonio. 2001. Mosquito: a Natural History of Our Most
Persistent and Deadly Foe. Faber
& Faber, London. |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}