File: <arthropodamed.htm> <Medical Index> <General Index> Site Description Glossary <Navigate
to Home>
ARTHROPODA
(Contact)
CLICK on underlined file
names and included illustrations to enlarge: The
Arthropoda is the most highly developed phylum of the invertebrates, and they
contain the greatest number of species.
Included are such widely recognized groups as the insects, spiders,
crabs and barnacles. General
characteristics are a segmentation that varies in different parts of the
body, often referred to as heteronomous
metamerism. There is a chitinous exoskeleton, which is
flexible but horny and tough. It is
always mixed with other materials such as proteins and fats. They all possess jointed appendages, one per segment, which are primitive and
never fully functional. The body
cavity is a haemocoel that is
not a true coelom. There are a dorsal
brain, ventral ganglia and nerve cord, and there is more nervous tissue in
the head than in the Annelida. They
have three body regions, which may be combined. The digestive tract is always of three parts: (1) foregut, (2) mid gut and (3) hind
gut. The foregut and hindgut are of
ectoderm derivation while the midgut is derived from mesoderm. Molting
is required as the cuticle is a non-growing part. ------------------------------------
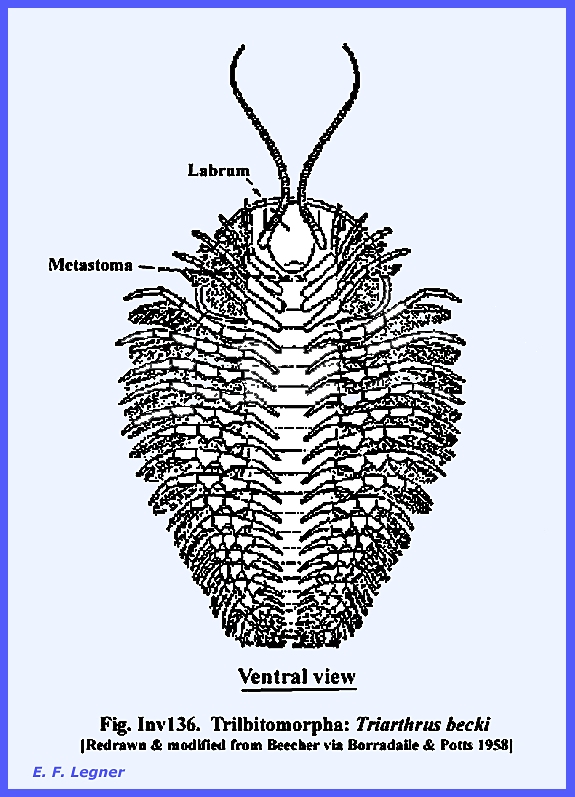
The Subphylum: Trilobitomorpha: Class: Trilobita – are an extinct group since the Paleozoic that is
studied because they were probably the most primitive of the Arthropoda. Their body had a central region with a
3-lobed lateral area on either side.
They had a head and trunk and were flattened dorso-ventrally. Compound eyes were present and one pair of
appendages occurred on every segment of the body save the first. Superficially they bore a close
resemblance to the Branchiopoda and also to the Brachyura crabs. Trilobites
were marine organisms and were numerous in the Cambrian and Silurian
eras. They were mostly quite small at
about 5 centimeters, but one fossil of Terataspis sp. reached about 0.7 meters in
length. The anterior somites of a
segmented trunk were movable on one another, while the most posterior
segments were united to form a tagma known as the pygidium. Their body
could be rolled up like that of an Isopod.
It is thought that most species lived at the bottom of the ocean in
shallow to moderately deep water.
However some species are believed to have been adapted to pelagic,
burrowing and deep-sea environments. It is
known that trilobites hatched as a larva, called the Protospis that
was a bit circular and consisted mostly of a head. See Inv136 for example.
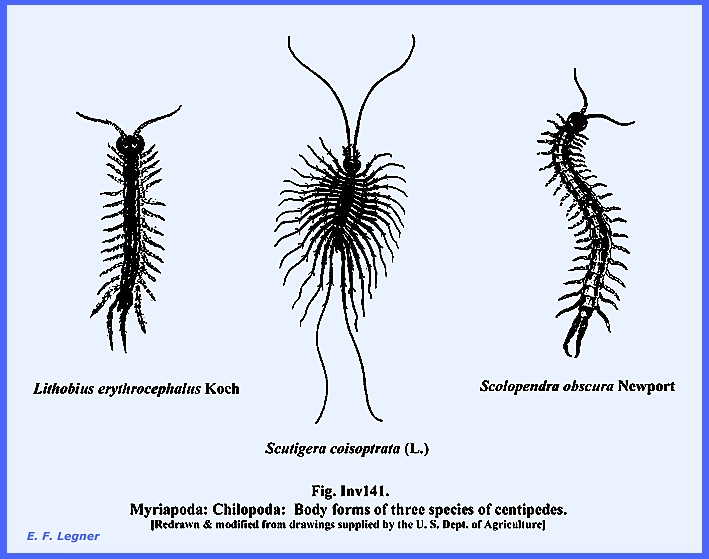
------------------------------------ Subphylum: Myriapoda, Class: Chilopoda includes the centipedes.
They are dorso-ventrally flattened.
Their body consists of a head and trunk but there is no thorax nor
abdomen. The head bears one pair of
antennae, one pair of mandibles, one pair of maxillipedes with poison
glands at the bases and ducts leading to pointed tips (Note: these are absent in the Diplopoda). There are two pairs of simple eyes called pseudocompound eyes. They have maxillae on the 1st and 2nd
segments. The trunk bears uniramous
appendages and there are 15 to 175 segments.
See examples at Inv141.
Body Wall -- This consists of a
cuticle, muscles and a haemocoel Digestive Tract -- A typical mouth
to anus arrangement. Circulatory System -- The heart is
tubular with one pair of ostia per segment.
The blood does not
carry oxygen Respiration -- The tracheae are
lined with ectoderm and cuticle, and heavy rings of cuticle line them. They branch out and ultimately reach all
tissues of the body. The blood does
not have an oxygen carrying function. Excretion -- Malpighian
tubules are long, thread-like and blind-ending tubules. They lie in the haemocoel and empty into
the digestive tract at the junction of the mid and hindguts. They extract nitrogenous wastes from the
blood. Nervous System -- This system is the
same as that found in the Crustacea. Reproduction -- The sexes are
separate. Genital organs are found at
the posterior end of the body and development is direct. Locomotion -- These animals are fast
movers. Long posterior legs are
sensory and used when moving backwards. Food & Digestion -- Chilopoda
are carnivorous and their food is paralyzed first by the maxillipedes. ------------------------------------ Subphylum:
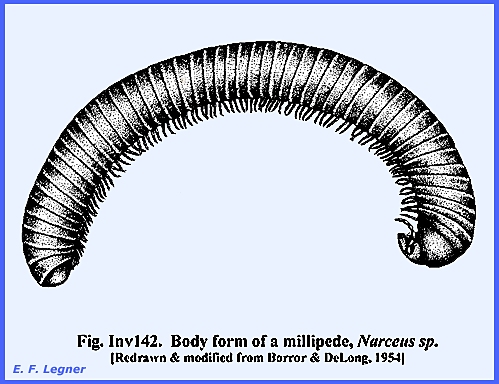
Myriopoda, Class: Diplopoda includes the millipedes.
These are cylindrical animals with a head and trunk that is the same
as in the Chilopoda. The head
appendages include antennae, mandibles, one pair of maxillae (instead of 2
pair as in the Chilopoda) and pseudocompound eyes on the head. The trunk has 25-100 or more segments with
each segment bearing two pair of appendages.
A fusion occurs between two segments all along the body except on the
first trunk segment. See example at Inv142.
Reproduction -- Genital ducts open
between the bases of the 2nd and 3rd legs.
This is anterior in contrast to the Chilopoda. Development -- it is essentially
direct except that the first larva has only three pairs of legs with
additions added later on. Food -- They are vegetarian animals
that move very slowly and are harmless. ------------------------------------ ============== |
|
Key References: <medvet.ref.htm> <Hexapoda> [Additional references may be found at: MELVYL Library] Akam, M. 2000. Arthropods:
Developmental diversity within a (super) phylum. Proceedings of the National
Academy of Sciences (USA) 97: 4438-4441. Averof, M. & M. Akam.
1995. Insect-crustacean
relationships: Insights from comparative developmental and molecular studies.
Philosophical Trans. of the
Roy. Soc., London B Biological Sciences 347: 293-303. Briggs, D.E.G., & R.A.
Fortey. 1989. The early radiation and relationships of
the major arthropod groups. Science 246 :241-243. Briggs, D. E. G., R. A. Fortey & M. A. Wills. 1993.
How big was the Cambrian explosion? A taxonomic and morphologic
comparison of Cambrian and Recent
arthropods. Pages 33-44 in Evolutionary Patterns and Processes (D. R.
Lees and D. Edwards, eds.) Linnean Society Symposium, Linnean Society of
London. Brusca, R. C. 2000. Unraveling
the history of arthropod diversification. Annals of the Missouri Botanical
Garden 87: 13-25. Brusca, R. C., & G. J. Brusca. 2003. Invertebrates. 2nd Edition. Sinauer Publ.,
Sunderland, Massachusetts. Chapman, A. D. 2009.
Numbers of Living Species in Australia and the World. 2nd edition.
Australian Govt, Dept. of Environ., Water, Heritage & Arts. Canberra. Chen, J. C., L. Ramsköld, & G.
Zhou. 1994. Evidence
for monophyly and arthropod affinity of cambrian giant predators. Science
263: 1304-1308. Cook, C. E., , Q. Yue, and M. Akam.
2005. Mitochondrial genomes
suggest that hexapods and crustaceans are mutually paraphyletic. Proc.Roy.
Soc Series B
272(1569): 1295-1304. Dohle, W. 1997. Are the insects more
closely related to the crustaceans than to the myriapods? Entomologia
Scandinavica Suppl. 51: 7-16. Edgecombe, G. D. (ed.). 1998.
Arthropod fossils and phylogeny. Columbia University Press, New York. Edgecombe, G. D. 1998.
Devonian terrestrial arthropods from Gondwana. Nature 394: 172-175. Edgecombe, G. D., G. Giribet, C. W. Dunn, A. Hejnol, R. M. Kristensen, R. C.
Neves, G. W. Rouse, K. Worsaae & M. V. Sørensen. 2011.
Higher-level metazoan relationships: Recent
progress & remaining questions. Organisms Diversity & Evolution
11(2): 151-172. Greenberg, B. 1973.
Flies and Disease. Vol. 2. Biology and Disease Transmission. Princeton
University Press, Princeton, NJ. 856 pp. Hall, R.D., Gerhardt R.
2009. Flies (Diptera). In Mullen Gr, Durden LA (eds.),
Med. & Vet. Entoml., 2nd Ed.. Elsevier, Burlington, MA. 637 pp. Harrison, I.H, G. A. Da Silva, M. Pitman & D. W.
Fleming, Vranjac A, Broome CV. 1989. Epidemiology and clinical spectrum of
Brazilian purpuric fever. Journal of Clinical Microbiology 27: 599-604. Hinkle, N. C., P. J. Scholl, D. E. Mock & W. B.
Warner. 2001. Research & extension needs for
integrated pest management for arthropods of veterinary importance. pp. 261-262. In Geden CJ,
Hogsette JA (editors), Proceedings of a workshop in Lincoln, Nebraska. 2nd Ed. 328 pp Janzen, J.-W. 2002. Arthropods in Baltic Amber. Ampyx-Verlag, Halle (Saale). Matheson, R. 1950.
Medical Entomology. Comstock
Publ. Co, Inc. 610 p. Müller, K. J. & D. Walossek.
1985. A remarkable arthropod fauna from the
Upper Cambrian "Orsten" of Sweden. Trans. Roy.Soc. Edinburgh: Earth
Sciences 76: 161-172. Peel, A. D., A. D. Chipman & M.
Akam. 2005. Arthropod segmentation: Beyond the Drosophila paradigm. Nature
Reviews Genetics 6(12): 905-916. Pisani, D., L.L. Polig, M. Lyons-Weiler & S. B. Hedges. 2004. The
colonization of land animals: molecular phylogeny and divergence times among
arthropods.
BMC Biol. 2(1). Service, M. 2008.
Medical Entomology For Students.
Cambridge Univ. Press. 289 p Legner, E. F. 1995. Biological control of Diptera of medical and veterinary
importance. J. Vector Ecology 20(1):
59-120. Legner, E. F. 2000.
Biological control of aquatic Diptera. p. 847-870.
Contributions to a Manual of Palaearctic Diptera, Vol. 1, Science Herald, Budapest. 978 p. Schram, F. R. & R. A. Jenner.
2001. The origin of Hexapoda: a crustacean
perspective. Pages 243-264 in Proceedings of the International Symposium on
the
Origin of the Hexapoda. T. Deuve, ed.
Annales de la Société entomologique de France 37. Shear, W. A. & J.
Kukalová-Peck. 1990. The ecology of paleozoic terrestrial
arthropods: the fossil evidence. Canadian Journal of Zoology 68: 1807–1834. Tudge, C. 2000. The
Variety of Life. Oxford: Oxford University Press. Waloszek, D. & J. A. Dunlop.
2002. A larval sea spider
(Arthropoda: Pycnogonida) from the Upper Cambrian 'Orsten' of Sweden, and the
phylogenetic position
of pycnogonids. Palaeontology 45: 421-446. Wheeler, W.C., P. Cartwright &
C.Y. Hayashi. 1993. Arthropod phylogeny: A combined approach. Cladistics 9: 1-39. |
FURTHER DETAIL = <Entomology>, <Insect Morphology>, <Identification Keys>