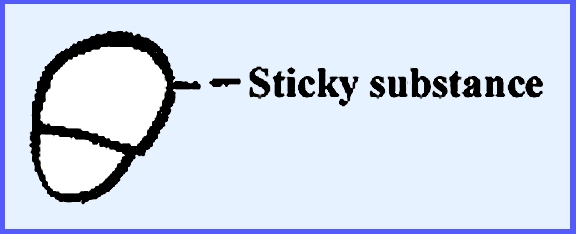File:
<ascomycetes.htm> <Index to Mycology> Pooled References <Glossary> Site
Description <Navigate to
Home>
Page 2
True Fungi (Eumycophyta1
Ascomycota (Ascomycetes,
Ascomycotina) -- Sac fungi
(Contact) Please CLICK on underlined
links & included illustrations for details Use Ctrl/F to search
for Subject Matter:
Tables Plates
Sample Examinations The order Sphaeriales
has a true peridium and
perithecial wall that is either brown or black in color. The texture is carbonaceous or
brittle. The ascus has rather thin
lateral walls and relatively a thick apical wall. Several members have perithecia scattered over a mycelial
mat. The grouping into families has
varied, but the present discussion includes eight: Chaetomiaceae, Fimetariaceae, Ophiostomataceae, Gnomoniaceae,
Diaporthaceae, Allantosphaeriaceae, Xylariaceae and Phyllachoraceae. The order might appropriately be
considered the "core group" of the Pyrenomycetes. Most of the Ascomycota that produce true
perithecia are included and eventually all such fungi might be incorporated
into the order. However, some of the
genera are being transferred to the order Pseudosphaeriales. The forms that will most likely retain an
enduring place in the Sphaeriales have typical flask-shaped perithecia. With
slender asci and paraphyses lining the lower portion of the fruiting body
cavity. The ascocarp wall is usually,
but not always, dark, and most often brittle or leathery in texture. The perithecia may be borne separately and
scattered on the mycelium, but they are sometimes produced in connection with
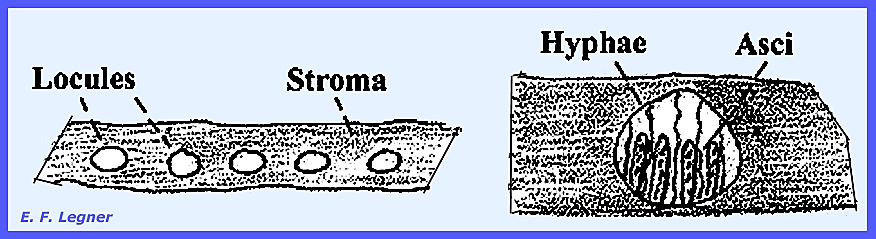
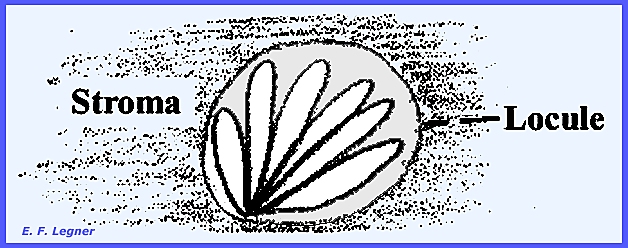
a stroma in which case they are usually closely clustered. In many genera the perithecia grow completely
imbedded in stromatic tissue, with only the ostioles protruding. Such perithecium-bearing stromata are
typically dark brown or black and sometimes carbonaceous in texture at
maturity. Many forms possess a
conidial stage like in the Hypocreales. The Family Chaetomiaceae is represented
by the Genus
Chaetomium. The ability to digest cellulose caused
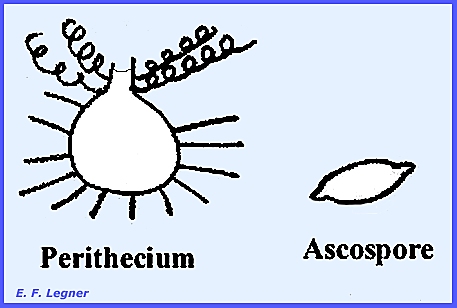
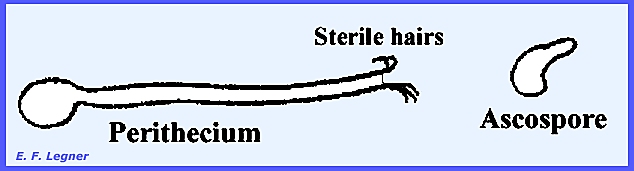
much difficulty with clothing during World War II. The perithecium is flask-shaped and sterile spine-like hairs
project from the flask. Coiled
appendages arise from the flask's neck.
The asci are round and the ascospores are citriform (lemon-shaped) and
they ooze out from slime at the neck.
The Family Fimetariaceae includes
the orange-colored bread molds. The
Genus Neurospora
is a common laboratory contaminant and it has a history of being a serious
pest in bakeries. The mycelium grows
very rapidly. The cells are
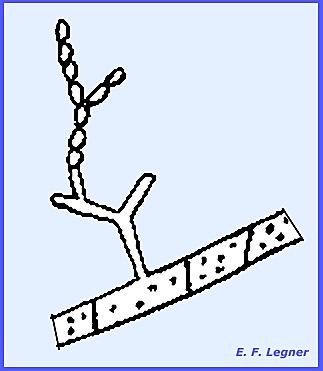
multinucleate and 1-4 days are required to complete the imperfect stage. The Monilia Imperfect Stage has branching conidiophores and a
multicellular, multinucleate mycelium.
There are branching chains of orange-colored conidia, which are often
called macroconidia. These are easily dislodged by air
currents.
The Perfect Stage is heterothallic
where one mating type (A or B) produces both sex organs, but neither will
mate with the same type.
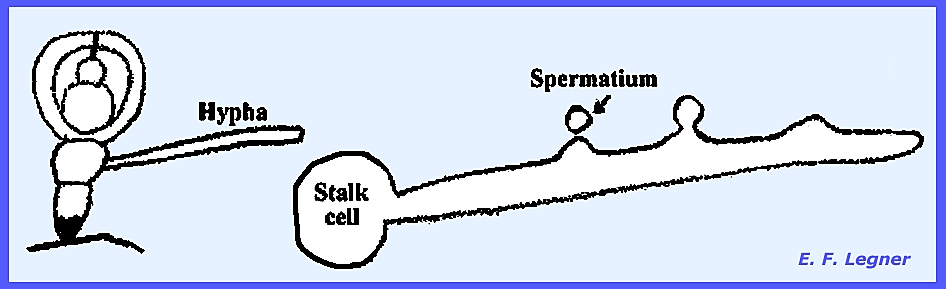
Spermatiophores are produced on all mycelia. These are flask-shaped structures, which form small,
uninucleate cells at the base (= spermatia or microconidia). Neurospora is exceptional in that
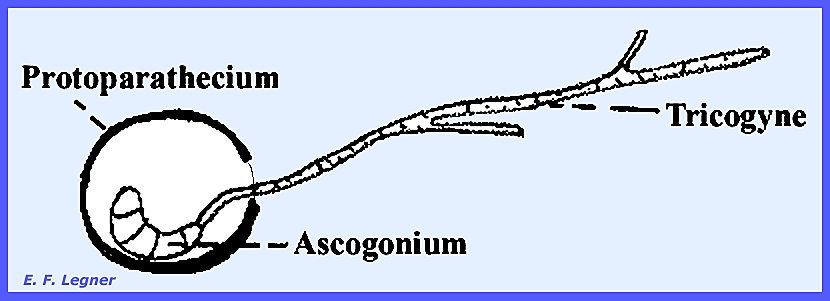
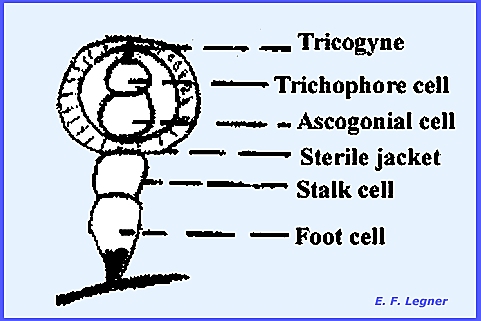
some of its spermatia may germinate directly. Protoperithecia are produced and contain a slightly coiled
ascogonium that is multicellular and surrounded by a weft of hyphae. From the apex of the ascogonium there
extends a relatively long tricoygyne, which may be branched and extended out some
distance. The tricogyne is narrower
than a hyphal cell, but has crosswalls and is multicellular.
The tricogyne grows toward the
spermatium if it is of the opposite mating type. Presumably the nucleus of the tricogyne after fusion with a
spermatium migrates down the tricogyne and joins with the ascogonium. Protoperithecia grow in size four hours
after mating, and perithecia are found with asci after crozier formation. Additional mating forms
exist. Mating with a macroconidium
may also be possible although the time for a reaction to the stimulus to show
is extended here. Mating between
hyphal cells is also possible.
Ascospores are forcibly discharged from elongated asci and only one
ascus at a time matures. Additional characteristics of Neurospora
are that the ascospores are small and the asexual cycle is completed in 1-2
days. The sexual cycle is completed
in 1-10 days. The mycelium is haploid
and all genetic markers appear and there is no masking by dominant
genes. This is a phototrophic genus
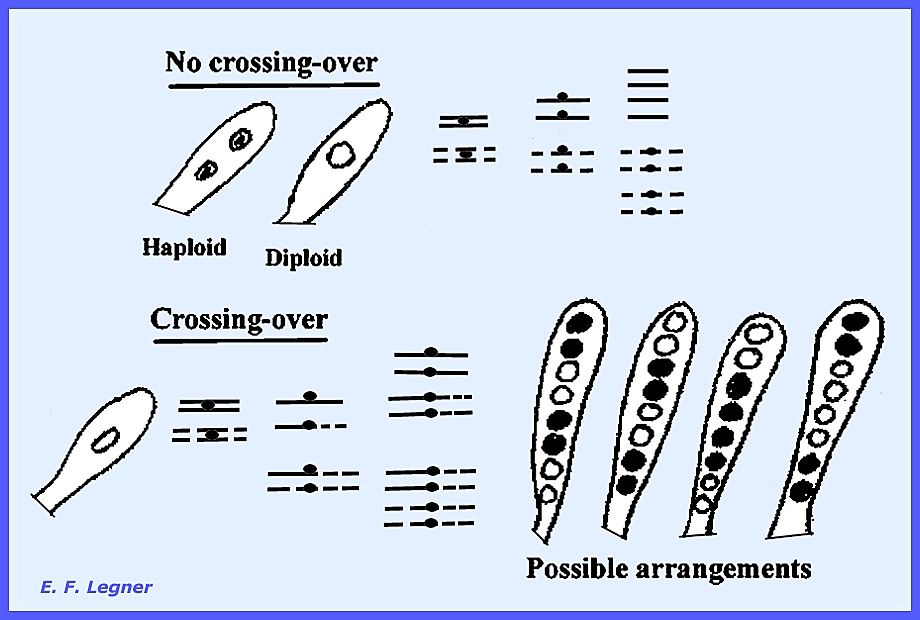
where all spores are thrown with equal force. ------------------------------------------- Neurospora sitophila and N. crassa have their asci formed in a
single row; the assumed position corresponds to the order in which division
took place (mitotic products). Thus,
it is possible to isolate single ascospores to determine where meiotic
processes occurred.
Neurospora tetrasperma was once thought to be homothallic
but it is actually facultatively homothallic. Two nuclei are enclosed in a common cytoplasm. Then, usually four ascospores are formed. Each ascus contains both mating
factors. Large ascospores are able to
give rise to functioning perithecia (homothallic), while small ascospores are
unable to do this and have only one nucleus each and are heterothallic.
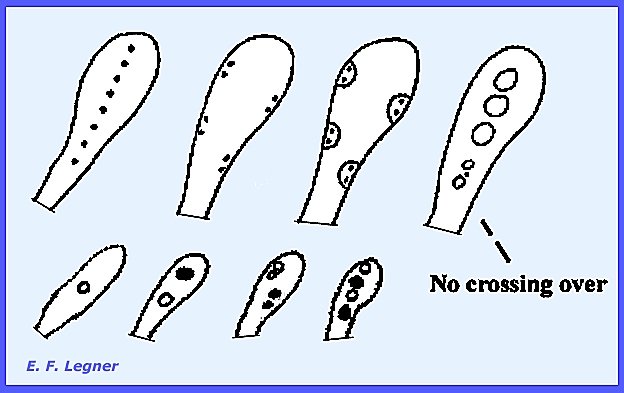
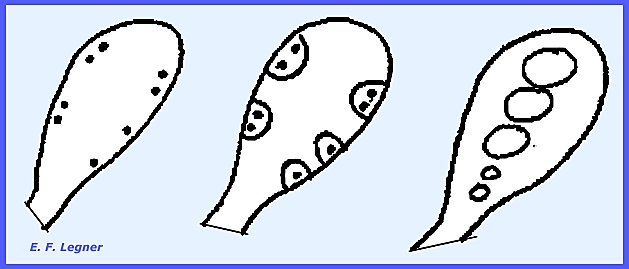
In most cases crossing-over does
take place.
When the nuclei do not lie close
enough together separate haploid ascospores are cut out.
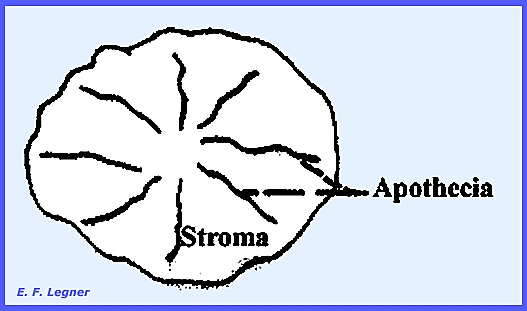
In the Family Xylariaceae
the Genus
Xylaria is sometimes referred to as
"Dead Man's Fingers." There is an u0right branched or simple
stroma. Perithecia line the outside
of the stroma.
The Genus Hypoxylon has a flat cushion-shaped stroma and the Genus Daldinia has
a round stroma with our without a stalk.
Characteristic concentric rings occur in the stroma, which are not
growth rings.
The Family Diaporthaceae is
represented here by Endothia parasitica.
These species reduced native American chestnuts to a non reproductive
state. This was a far superior
species than those occurring in Europe.
Invading from the Orient it was first observed in America in 1910 in
New York state. The mycelium of this
fungus kills the chestnut cambium.
The mycelium forms a stroma just under the surface of the bark. Perithecia are embedded, necks only, at
the base of the stroma. These necks
are very long and extend up through the stroma.
The Family Allantosphaeriaceae
differs from the Diaporthaceae by having asci with a long, tapering,
persistent stalk and they form a definite hymenial layer in the
perithecium. Paraphyses are formed
but they gelatinize by the time the perithecium is mature. The family gets its name from the
ascospores that are usually sausage-shaped (allantoid). They are one to many-celled and they are
brown in color when occurring in a mass.
Most species are saprobic, inhabiting dead bark or wood, with a few
species being also parasitic on some plant species. In the Family Phyllachoraceae the
Genus Phyllachora
causes "Tar Spot"
of grasses. In some species the perithecium
practically fills the space between the cuticle and the epidermis. Stromatic layers are formed between the
necks and bases of the perithecia.
The Family Ophiostomataceae includes some severe
plant pathogens. The Genus Ceratocystis causes "Blue
Stain" of wood. One notorious species, Ceratocystis
ulmi, causes "Dutch Elm Disease"
of American elm. It was first detected
in North America in 1931. By the year
2000 over 98 percent of American elms were destroyed. The only survivors occur on the large
islands of Northeast Canada. The fungus is heterothallic and the scattered
perithecia have extremely long necks.
Ascospores are slightly curved and pushed outside in a gelatinous
mass. The asci disintegrate early so
that spores lie free in the perithecial wall.
In another Genus Ustulina a layer of perithecia occurs on a convoluted stroma.
The Family Gnomoniaceae have
their perithecia buried in the substratum and each of them is provided with a
prominent beak that protrudes from the surface and serves as an exit for the
ascospores. The ascal walls of some
species are much thickened at the apex of the ascus to form a narrow canal in
the center through which the ascospores pass while being discharged. There are no paraphyses in the mature
perithecia. Gnomonia leptostyle causes
anthracnose of walnut and
related plants. Gnomonia
veneta causes sycamore anthracnose and G. ulmea causes leaf
spot of elm.
Gnomonia erythrostoma is the cause of cherry
leaf scorch. The conidia of most species are produced
in acervuli. However, G.
fragariae on strawberry bears conidia in pycnidia. ----------------------------- Please refer to the
following plates for characteristic structures and Life Cycles in the Sphaeriales: Ascomycota: Euascomycetes: Sphaeriales Plate 128 = Sphaeriales: Ascocarp diagram. Plate
129 = Chaetomium
sp.: Ascospore maturation. Plate
130 = Life Cycle -- Neurospora sitophila. Plate
131 = Structures of Ophiostoma fimbriatum. Plate
132 = Structures of Gnomonia ulmea: Perithecium
& Ascus. Plate
133 = Structures of Gnomonia fragariae. Plate
134 = Structures of Phyllachora graminis:
Stroma & perithecia. Plate 205 = Life Cycle -- Pyrenomycetes: Sphaeriales: Neurospora
sitophila Plate 206 = Diagnostic Characters: Pyrenomycetes: Sphaeriales: Ceratocystis, Chaetomium, Daldinia, Neurospora, Phyllachora, Ustulina, Xylaria Plate
209 = Diagnostic Characters -- Pyrenomycetes: Sphaeriales: Chaetomium chamalodes & Neurospora
tetrasperma Plate
210 = Example Structures -- Pyrenomycetes: Sphaeriales: Ceratocystis sp., Ceratocystis ulmi, Daldinia
sp.,Endothia parasitica, Hypoxylon sp., Neurospora tetrasperma, Phyllachora sp.,
Ustulina sp., Xylaria cornu-damae, Xylaria spp. ----------------------------- The order Pseudosphaeriales is a rapidly growing
order, for many forms are being transferred to this group from other areas of
the Pyrenomycetes. They are mostly tropical
species and ascocarps that are not true perithecia, and thus they are
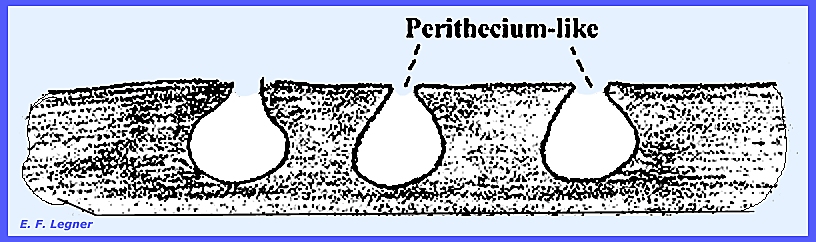
referred to as "perithecia-like" or "ascostroma." They become exposed to the external
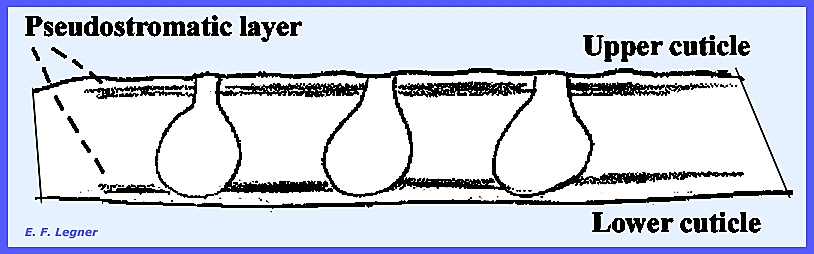
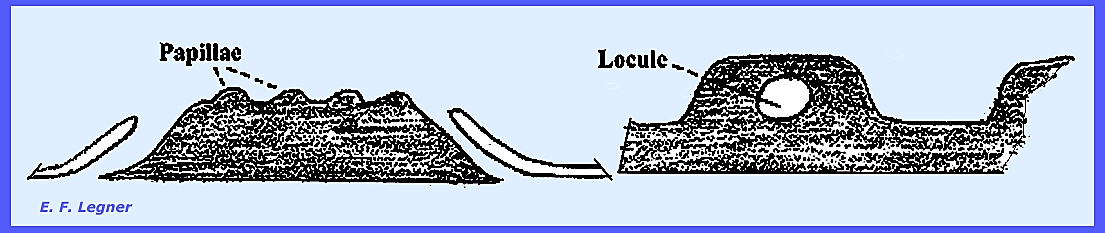
environment by pushing through the host cuticle. Locules
are formed in the stroma and hyphae grow from the top to the base
of the locule. They usually persist
through maturation and they are called pseudoparaphyses, and no
perithecial peridium is formed here.
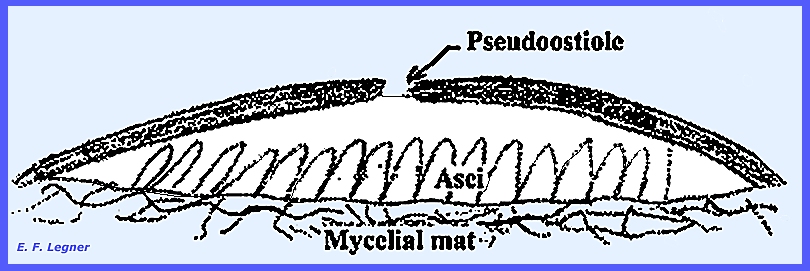
The ascus wall is uniformly
thickened and is actually composed of two layers with no pore at the
apex. During release of ascospores
the outer wall ruptures and the inner wall balloons-out.
Dibotryon morbosum causes
"Black Knot" disease on plum
and cherry. The fungus invades the
young, elongating shoots of cherry in the spring. The cambial layer of the host is reached by autumn but there is
no obvious damage. The mycelium
overwinters; and during the second growing season stimulates the cambium of
the host to divide rapidly. Also the
ray cells are enclarged. This
hypertrophy breaks up the cambium.
There is also a differentiation of host cells produces a mass that is
difficult to distinguish between wood and bark. The mycelium constructs a stroma underneath the hard bark, in
the vicinity of the cork cambium. The
fungus stroma then ruptures the hard bark of the host.
Many conidia are produced on the
surface of the stroma, and there is a Hormodendrum Imperfect Stage in
early summer. After the imperfect
stage the stroma pushes up a number of papillae on its surface.
Ascogenous hyphae develop in
locules of the pipillae and an opening or pseudoostiole forms in the
second spring. Asci deliquesce and
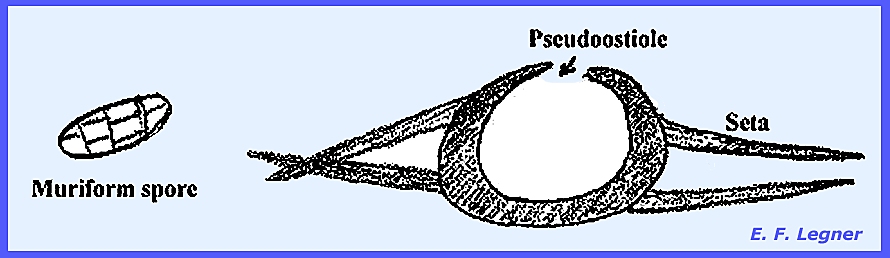
ascospores are 2-celled being distributed via the psuedoostiolein the locule. Pyrenophora teres causes "Net
Blotch" of barley. Ascostromata are formed and a
pseudoostiole allows for the escape of spores. These are muriform, and setae are found on the sides of the
ascostroma.
Pleospora sp. are similar to
Pyrenospora teres except that there are no setae. Venturia inaequalis causes
the "Apple Scab" disease. Here the mycelium is subcuticular (between
the cuticle and the epidermal cells).
It does not penetrate the host nor produce haustoria and possesses a
curious nutrition that is not fully understood. The mycelium produces a number of upright, unbranched
conidiophores; each being terminated by a single conidium. This is the Fusicladium Imperfect Stage (like
small candles)
After the conidium is released
another forms in its place. The
parasitic stage is also the Imperfect Stage here. The Saprophytic Stage of V.
inaequalis occurs on dead material in winter. A stroma forms during winter and the ascogonium is embedded
therein a forms a short tricogyne. An
antheridium fuses with the tricogyne and its nucleus migrates down a tube to
the ascogonium. Although both sex
organs are produced on the same thallus, there are compatibility types
present. Asci mature by spring and
are found in locules in the stroma.
Ascospores are 2-celled with one cell larger than the other and they
are initiated in the fallen leaves in autumn but do not mature until the
following spring. Spores are
discharged from the psuedoostiole.
----------------------------- Please refer to the
following plates for characteristic structures and Life Cycles in the Pseudosphaeriales: Ascomycota: Euascomycetes: Pseudosphaeriales Plate 139 = Life Cycle -- Mycosphaerella tulipiferae. Plate
140 = Structures of Guignardia bidwellii. Plate
141 = Life Cycle -- Venturia inaequalis. Plate 211 = Life Cycle -- Pyrenomycetes:
Pseudosphaeriales: Venturia inaequalis Plate 212 = Example Structures --
Pyrenomycetes: Pseudosphaeriales:
Dibotryon, Pyrenophora, Pleospora, Venturia Plate 214 = Example Structures #2: Pyrenomycetes: Pseudosphaeriales, Dothideales, Hemisphaeriales,
Laboulbeniales, Hysteriales. Plate 215 = Example Structures #3: Pyrenomycetes: Pseudosphaeriales, Dothideales,
Hemisphaeriales,
Laboulbeniales, Hysteriales. ----------------------------- The order Dothidiales is mainly
tropical and has neither paraphyses nor pseudoparaphyses. A locule is digested in the stroma and
asci begin to grow in the locule before the latter is mature (unlike
Pseudosphaeriales). All asci appear
to originate at a point giving a form like a fan at maturity.
Ascospores are released in the
same manner as in the Pseudosphaeriales.
A pseudoostiole allows for the escape of spores to the external
environment. Most species are
tropical. Mycosphaerella sp. has only one locule per
stroma. Cymadothea, causing "Clover
Sooty Blotch" has more than
one locule per stroma and there is a Polythrincium Imperfect Stage. The Perfect Stage matures on overwintered
leaves, but is very rare. Dothidea
collecta is
multiloculate. Ascogonia are embedded
in the ascostroma and a locule dissolves around each ascogonium. A pseudoostiole is present.
----------------------------- Please refer to the
following plates for characteristic structures and Life Cycles in the Dothideales: Ascomycota: Euascomycetes:
Dothideales Plate
138 = Dothideaceous stroma: Typical structure. Plate 213 = Example Structures --
Pyrenomycetes: Dothidiales,
Hemisphaeriales, Laboulbeniales, Hysteriales Plate 214 = Example Structures #2: Pyrenomycetes: Pseudosphaeriales, Dothideales, Hemisphaeriales,
Laboulbeniales, Hysteriales. Plate 215 = Example Structures #3: Pyrenomycetes: Pseudosphaeriales, Dothideales,
Hemisphaeriales,
Laboulbeniales, Hysteriales. ----------------------------- The order Hemisphaeriales
(or Microthyriales) is represented by the
Genus Asterina. There are many tropical species. These are
leaf parasites that bear a hemispherical ascostromata. The ascocarp is superficial on the host or
sometimes subcuticular. The top
portion of the stroma is clearly defined and the bottom is a mat of loose
hyphae. The asci come to lie beneath a shield-like cover that opens by
irregular spitting or may form a pseudoostiole. Pseudoparaphyses may be present.
----------------------------- Please refer to the
following plates for characteristic structures and Life Cycles in the
Hemisphaeriales: Ascomycota: Euascomycetes:
Hemisphaeriales Plate
142 = Structures of Hemisphaeriales: Microthyrium
microscopieum. Plate 213 = Example Structures --
Pyrenomycetes: Dothidiales,
Hemisphaeriales, Laboulbeniales, Hysteriales ----------------------------- The order Laboulbeniales includes
nonpathogenic parasites of insects.
There are over 100 genera and thousands of species have been
described. The species discriminate
the site of the insect body they will attack. Many species are extremely minute and are composed of
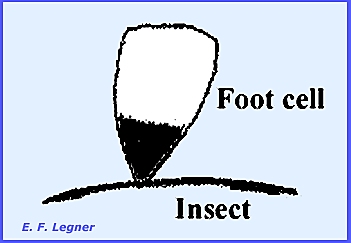
relatively few cells and some species are dioecious. The thallus is non-mycelial and attaches
to the host by means of a shield-shaped foot cell with a black pigment at its
apex.
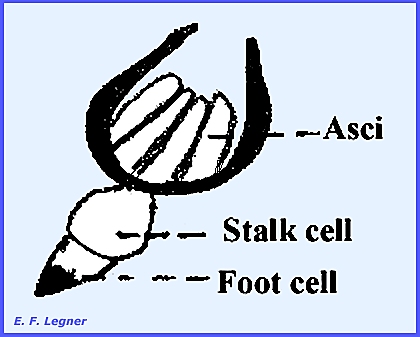
Rhizoids have been observed to
enter the body of the host. A stalk
cell lies on top of the foot cell, which bears a jacket of sterile cells.
Spermatia are produced in the
hypha that extends from the stalk cell.
The spermatium joins with a
tricogyne and its nucleus fuses with the ascogonium nucleus. A jacket of sterile cells enlarges.
Asci are developed within and
generally have four ascospores (others disintegrate). These are forcibly ejected. Male and female fungi generally occur
close together on the host, and the ascospores are 2-celled. The larger cell, which is equipped with a
sticky substance, goes to the foot cell and the smaller one gives rise to the
rest of the thallus.
----------------------------- Please refer to the
following plates for characteristic structures and Life Cycles in the Laboulbeniales. Ascomycota: Euascomycetes:
Laboulbeniales Plate
127 = Structures of Laboulbenia formicarum. Plate 213 = Example Structures --
Pyrenomycetes: Dothidiales,
Hemisphaeriales, Laboulbeniales, Hysteriales Plate 214 = Example Structures #2: Pyrenomycetes: Pseudosphaeriales, Dothideales,
Hemisphaeriales,
Laboulbeniales, Hysteriales. Plate 215 = Example Structures #3: Pyrenomycetes: Pseudosphaeriales, Dothideales,
Hemisphaeriales,
Laboulbeniales, Hysteriales. ----------------------------- The order Hysteriales
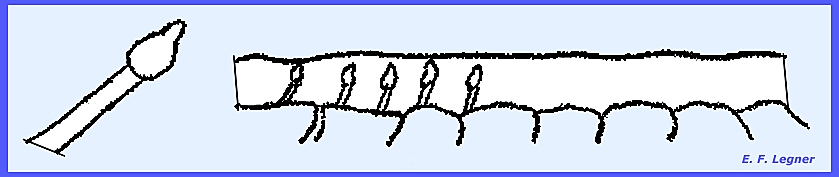
forms elongated
ascostromata, which are black, hard and carbonaceous. They may be closely packed on wood, giving
the appearance of charred wood.
An ostiole opens in an elongated slit
called a Hysterothecium. The asci are of the Pseudosphaeriales type
and pseudoparaphyses may be present. ----------------------------- Please refer to the
following plates for characteristic structures and Life Cycles in the Hysteriales: Ascomycota: Euascomycetes:
Hysteriales Plate
143 = Ascocarp & ascus of Hysteriales: Glonium
sp. Plate 213 = Example Structures --
Pyrenomycetes: Dothidiales,
Hemisphaeriales, Laboulbeniales, Hysteriales Plate 214 = Example Structures #2: Pyrenomycetes: Pseudosphaeriales, Dothideales, Hemisphaeriales,
Laboulbeniales, Hysteriales. Plate 215 = Example Structures #3: Pyrenomycetes: Pseudosphaeriales, Dothideales,
Hemisphaeriales,
Laboulbeniales, Hysteriales. ----------------------------- The Sub-Class Euascomycetes, Series Discomycetes are the "cup
fungi," a group with
over 11,000 species. The ascocarp is
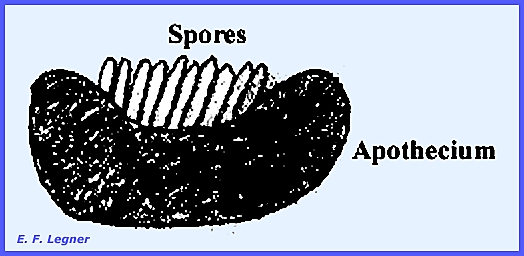
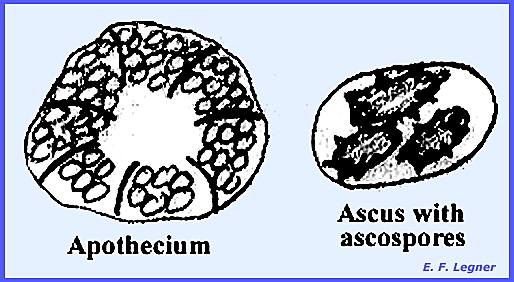
an apothecium. The hymenium is always
exposed and cup-shaped.
There are two Sub-Series: Inoperculatae and Operculatae. The Inoperculatae are mainly parasites
that do not have an operculum or "lid" on the ascus. Most species have a conidial stage and the
fruiting bodies are quite small. They
are usually not coprophilous and the majority produce spermatia. The Operculatae are not usually parasites
and they do possess an operculum (boudier).
Most
species are without a conidial stage and they usually have large fruiting
bodies. A large number are
coprophilous and not one produces spermatia.
Their ascospores are not elongated. The Inoperculatae are here
represented by two orders: Helotiales
and Lecanorales. The order Helotiales is the larger of the
two orders of Inoperculatae. They hve
either cup or disc-shaped apothecia with asci only slightly thickened at the
apex. The ascospores are round,
elliptical or elongated but rarely thread-like. Many of the Helotiales live as soil saprophytes on dead wood or
dung or on other organic matter from which they derive nourishment. Some species are very serious parasites on
plants, causing various rots of stone fruits and diseases of vegetable
crops. Several families and genera
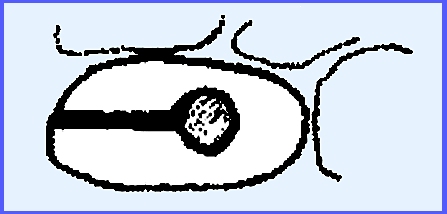
will be discussed to represent this order. ----------------------------- In the family Hypodermataceae both Hypodermella
and Lophodermium
incite "Needle-Cast Disease"
of conifers. The fruiting body is
boat-shaped with a slit down the middle.
It shows a resemblance to the ascocarps of the Hysteriales, although
it is generally much smaller and less carbonaceous. They are really a
transitional group between the Pyrenomycetes and Discomycetes and this
fruiting body is like a hysterothecium.
The asci are not double-walled
here, a spermatial-type of fertilization occurs and conidia are quite rare. Rhytisma acerinum causes
"Tar Spot Diseases." There is conspicuous black stroma on
plants, which is produced close to the surface or directly in the epidermis
(= superficial stromata). Ascocarps
develop within the stroma and spermatia are produced in the initiation of the
sexual process. The ascocarps are in
the stroma and radiate out from the center.
They stand out as prominent lines on the stroma. The hymenium is broader than in previous
forms.
Cryptomyces pteridis is essentially the same as Rhytisma
acerinum but is parasitic on bracken fern in North America. There is a conspicuous black
stromata. Several elongated
apothecia, more or less radiately arranged, develop in each stroma, maturing
late in spring on the dead overwintered leaves lying on the ground. The fungus in its parasitic phase has a
conspicuous intercellular mycelium with haustoria, and produces large
quantities of long, slender, curved spores in acervuli. These break through the epidermis (the Cylindrosporium Imperfect Stage). If the infection is heavy the leaves
usually turn yellow and fall off. ----------------------------- The family Mollisiaceae is represented here by Higginsia hiemalis (Cocomyces),
which causes "Cherry Leaf Spot"
disease. There is a rapidly developing
septate mycelium in intercellular spaces that produce haustoria with long
stalks and a bulbous tip. There is a
prominent conidial stage.
In the Cylindrosporium Imperfect Stage an aggregation
of hyphae forms in a little bed under the epidermis. Conidiophores that are
produced on top produce elongated conidia, which are either one- or
two-celled or slightly curved.
This mass of protruding conidia
breaks the cuticle and is conspicuous (= acervulus). In the Sexual Stage apothecia form
on overwintered leaves on the orchard floor.
The sexual process is initiated on the leaves still on the tree but
late in the season. Tiny cells are
formed at the base of the stroma, which are spermatiophores. They seem to be derived from
conidiophores. These produce
spermatia.
Down in the leaf there is a
massing of hyphae that enlarges (= ascocarp initial). Coiled hyphae are produced on the mass,
which develop into a tricogyne, which migrates to the surface. Spermatia attach themselves to the
tricogyne.
The stroma enlarges over winter and
asci develop. They differ from the
Pseudosphaeriales in that there is no double wall and they occur with
paraphyses on the hymenium.
Stromatic tissue cracks open like
a lid, exposing the hymenium (= apothecium is produced from a stroma the top of which breaks
off). Ascospores are straight and
never more than 2-celled (remember that conidia may be 2- or 3-celled).
Diplocarpon rosae causes "Black
Spot Disease" of rose. Most
characters are like Higginsia but the mycelium is subcuticular and not
as deep as Higginsia. There is
an Actinonema
Imperfect Stage. The
conidia and ascospores are similar.
Febaea sp. is mostly like Diplocarpon
but there is a different Imperfect Stage.
The ascocarp may, in a few cases, develop on living tissue of the
host. Mollisia dahnii grows as a parasite on the
stems and leaves of Potentilla, a wild rosaceous plant related to the
strawberry. The apothecia are matured
on the living host. ----------------------------- The family Helotiaceae is
represented here by several genera. Stamnaria americana is of interest as one
of the very few fungi that attack horsetails (Equisetum spp.). In autumn numerous apothecia develop and
mature on the live stems of the host..
It infects stems and produces ascocarps in autumn on living stems. They possess a rather prominent stalk.
The ascocarp is made up of
obviously interwoven hyphae (= prosenchyma). The Mollisiaceae
had a stroma, which is a group of very closely oppressed hyphae called a pseudoparenchyma. Chlorosplenium aeruginosa
causes "Green Stain" disease, which
is easily observed with the naked eye.
It is bluish-green in color and found on oak wood. The green stain is highly prized in
Europe. Ascocarps are produced from
mid-summer to the end of October. The
hyphae contain a green pigment and their color is imparted to the
substrate. The apothecia are also
green. They are stalked, cup-shaped
to funnel-form at the apex where asci are borne, and often asymmetrical. ----------------------------- In the family Sclerotineaceae apothecia are typically
long-stalked with funnel-like tips.
They always arise from a slerotium or pseudosclerotium. Most species are parasitic and there is
definite spermatial fertilization.
Monilinia fructicola causes
"Brown Rot" of stone
fruits. This is a highly economically
important fungus that attacks fruit at the ripening stage and may develop in
transit. The Monilia Imperfect Stage is similar to Neurospora. Apothecia originate from a
pseudosclerotium. Tufts of conidia are
formed on the host surface, the fruit becoming mummified (it is not a soft
rot). The mummy bears the
pseudosclerotium. Partially buried
mummies on an orchard floor give rise to stalked apothecia. Spores are commonly one-celled and
hyaline. Sclerotinia
sp. never has a Monilia Imperfect Stage and apothecia
originate from a true sclerotium. ----------------------------- The family Geoglossaceae are known as the "Earth
Tongues." All are saprophytes that grow out of the
ground. Apothecia are never cup or
saucer-shaped and there is no conidial stage. Instead apothecia are tongue-, cup or spatula-shaped Details of
the sexual stage are not well understood.. Geoglossum sp. has a
hymenium that spreads on all sides of the apothecium. Ascospores are distinctive.
Leotia lubrica. has a white stalk
where the upper portion is convoluted into a dark green cup-like area. The fungal body is very slippery. ----------------------------- Please refer to the
following plates for characteristic structures and Life Cycles in the
Helotiales: Ascomycota: Euascomycetes:
Helotiales Plate
144 = Section thru' Apothecium diagram. Plate
145 = Life Cycle -- Rhytisma acerinum. Plate
146 = Life Cycle -- Monilinia fructicola. Plate
147 = Geoglossaceae structures: Geoglossum
ophioglassoides, Spathularia clavata, Leotia
gelatinosa & Cudonia circinans. Plate 216 = Life Cycle -- Discomycetes:
Inoperculatae: Helotiales: Hypodermataceae:
Rhytisma acerinum Plate 217 = Diagnostic Characters --
Discomycetes: Inoperculatae: Helotiales: Geoglossaceae, Helotiaceae,
Hypodermataceae, Mollisiaceae, Sclerotineaceae Plate 218 = Example Structures --
Discomycetes: Inoperculatae: Helotiales:
Hypodermataceae Plate 219 = Example Structures --
Discomycetes: Inoperculatae: Helotiales:
Helotiaceae, Mollisiaceae Plate 220 = Example Structures --
Discomycetes: Inoperculatae: Helotiales:
Helotiaceae, Sclerotineaceae,
Geoglossaceae ----------------------------- The order Lecanorales includes
over 8,000 species, all of which are parasites on algae forming lichen
associations. This is
considered to be a symbiosis: the
alga derives protection from the fungus, which in turn derives nourishment
from the alga. In actuality the
fungus makes a slave out of the alga = Helotism. Lichens have actually been given
binomials. Other fungus groups that
together with algae form lichens are in the Pyrenomycetes (about 2,000
species in Sphaeriales & Pseudosphaeriales) and in the Agaricales. About 18 families have been
recognized in the Lecanorales. They
are widely distributed in nature from the poles to the tropics. They are especially abundant in the Arctic,
where they comprise the principal vegetation, and they are important there
for forage of musk oxen and reindeer and have been harvested by humans for
their herds. They can live on bare
rock; withstand long periods of desiccation, severe cold and severe
heat. They do not die in the winter
and they may dry down to brittleness, but can reabsorb water. Classification of Lecanorales is
made on the thallus form. Foliose =
like a sheet; Fruticose = upright and profusely branched or those that hand
from trees, and Crustose = thallus closely oppressed to the substrate. Lichens are anchored to the substrate and
they weather the rock (the first step in soil formation). They may break open the rock by sending hyphae
into the crevices and acid secreted digests the rock. They are extremely sensitive to noxious
gases and are rarely found in urban areas. The kinds of algae parasitized are
unicellular green algae, blue-green algae (Chlorella) and filamentous green
algae. Algae may multi80ly by fission
when in association with the fungus. Parmelia sp. forms a compact
layer of interwoven hyphae on two sides (= pseudoepidermal layer). A loose system of hyphal threads extends
between these. Rhizines are produced at the
base, which serve for anchorage and absorption of some water and minerals
from the substrates. The algae occur
near the surface surrounded by a loose mass of hyphae (they are always
embedded in the fungus thallus). The
alga is protected from direct rays of the sun as the fungal covering produces
the blue-green color of lichen. This
also lowers the risk of desiccation.
Hyphae derive food from the alga in two ways: (1) by hyphal contact and (2) by small
haustoria.
Propagation of the fungus usually
occurs by braking off of a portion of the thallus through the agency of wind,
etc. This process is known as Multiplication
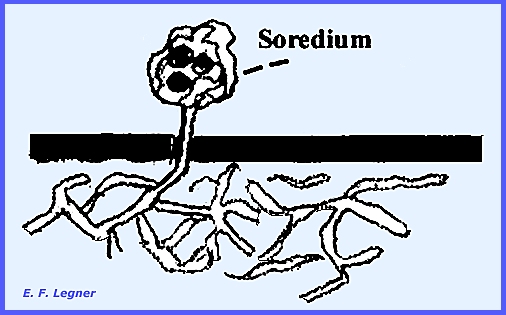
by Fragmentation. A soredium occurs
where several algae are suspended by some fungal hyphae above the
pseudoepidermal layer.
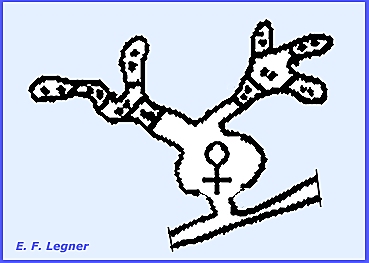
During ascospore formation an
ascogonium that is produced down in the hyphal mass, which sends up a
tricogyne.
A spermogonium is produced with an ostiole protruding from the
pseudoepidermal layer.
After the spermatia attach to the
tricogynes, the male nucleus migrates down the tube of the tricogyne and
fuses with the female nucleus in the ascogonium. Ascogenous hyphae form croziers and these produce asci. In the meantime the apothecium is formed
around the ascogonium. Ascospores are
forcibly discharged. Apothecia form
on the surface, and algae may or may not be carried along into the ascocarp.
Ascospores will not germinate
unless they alight on a specific alga, which is one in a million! Lichens are an extremely important
group of organisms. They colonize
barren lands and serve as forage for animals. They may supply food for humans in emergencies, but most are
toxic and boiling or leaching is required.
They have been used in the production of purple dyes and in the
perfume industry. They can cause
extensive damage to stained glass windows in cathedrals and indirect damage
to trees by harboring insects.
----------------------------- Please refer to the
following plate for characteristic structures in the Lecanorales: Ascomycota: Euascomycetes: Lecanorales Plate
221 = Example Structures -- Discomycetes: Inoperculatae: Lecanorales ----------------------------- The order Pezizales
and two families Pezizaceae
and Helvellaceae here represent
the Operculatae. Almost all Operculatae are saprophytes on dead
wood, leaves, dung, etc. They have an
operculum or lid on the ascus, and very few forms produce conidia. Fruiting bodies are considerably larger
than the Inoperculatae. There are no
spermatia. Ascospores are one-celled,
never markedly elongated and often ornately sculptured. The order Pezizales may
have cup or saucer-shaped apothecia and stalked apothecia depending on the
family. The family Pezizaceae
or cup
fungi is
represented here by several genera with different characteristics. It is a relatively large group of
saprophytic forms that grow on old wood, rich soil, dung, etc. Their apothecia are typically cup-shaped
or discoid, and may be stalked or sessile.
There is a wide range in size, color and other details. Some species are heterothallic. The Genus Peziza has an apothecium
that may be one foot in diameter. The
Genus Aleuria
has reticulate sculpturing on the ascospores. The Genus Urnula is the earliest to appear of the fleshy fungi in
North America. They always occur on
buried wood and they have black spores and ascocarps.
Paxina and Plectania
have a well-developed stalk and diagramatic cup. Ascobolus has many coprophilous species. Apothecia are quite thick and have purple spores.
Pyronema grows where soil
has been heated, e.g., around old campfire sites, in greenhouses, after a
forest fire, etc. Sexual Reproduction
in The Operculatae Many forms in the Pezizaceae have
been studied, but not a single member in the whole order produces
spermatia. An example is Pyronema
where there is development of branches from certain hyphae that are swollen
at the tip and delimited by a cross-wall.
Many nuclei pass from the antheridium into the ascogonium. The nuclei then pair in the ascogonium.
Paired nuclei move out into
ascogenous hyphae.
At the same time that ascogenous
hyphae are produced, sterile hyphe and paraphyses grow from beneath the
ascogonium. Thus the stalk and excipulum are
formed. Another example is Ascobolus that contains a considerable number of coprophilous
species. They have small apothecia,
1-2 mm. in diameter. Ascospores are
large with purple spores. During
sexual reproduction the ascogonium occurs with an antheridium. These ascogonia may be coiled and
tapered. Ascogenous hyphae form from
the coil after fertilization.
----------------------------- The
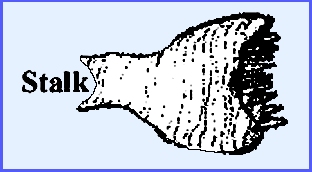
family Helvellaceae is
the saddle fungi all of which are
saprophytes. The apothecium is
always stalked and never cup-like.
The Genus Helvella
has the following saddle shape:
The Genus Morchella has a hymenium that lines all the depressions. All are edible and highly prized. The mycelium persists in the ground from
year to year.
In the Genus Verpa, the ascospores vary
from globular to elliptical, but never elongated nor mullticellular. Spores may be sculptured (a condition is
essentially nonexistent in the Inoperculatae). ----------------------------- Please refer to the
following plates for characteristic structures in the Pezizales: Ascomycota: Euascomycetes: Pezizales Plate 148 = Ascocarp & ascus: Elvella
crispa. Plate 222 = Example Structures --
Discomycetes: Operculatae: Pezizales & Tuberales ----------------------------- The order Tuberales includes
the truffles,
which are highly coveted for their flavor.
There are more than 200 known species of which about 62 have been
found in America, mainly in California.
The fruiting bodies are mostly subterranean, and those of certain
species in the genus Tuber are the most highly prized. The fruiting bodies are mostly closed
structures and do not resemble apothecia.
Most species do not have an ascal opening. The ascocarps range in size from smaller than a pea to several
centimeters in diameter. They are
somewhat globose, often fleshy structures with a smooth or irregular
surface. The asci line one or more
chambers, which open to the outside; or the ascocarp may be essentially
solid, closed and indehiscent, its asci appearing to be scattered and
imbedded in sterile tissue. The
fructification of the Tuberales has been interpreted as a modified
apothecium, and this has directed its classification into the discomycete
group and close to the Pezizaceae. The Genus Tuber has mycorhizal forms on oak roots. They occur all over the world, but are
commercially harvested in France, Italy and Spain. They are also common along the West Coast of North
America. Truffle hunters use dogs and
pigs to hunt down the fungi, which are buried several inches below the
ground. Ascocarps may be up to two
inches in diameter. Veins separate
the globular scattered asci. This is
actually a very highly evolved apothecium.
The cup is convoluted into a system of folds in evolution; asci are
typically operculate. Asci are also
globular and the spores are sculptured, 1-celled and short. Typically there are 2-5 spores per ascus.
The Genus Genea is more primitive than Tuber and contains a small
ostiole.
The Genus Pseudobasamia causes "Calves
Brains Disease." The apothecium is quite small (1/2 the
size of a pea), and it is produced in convoluted masses. These are pests of the mushroom industry
as they inhibit the growth of commercial varieties. Spores of the Tuberales are
generally distributed by animals, which dig them up for nourishment, the
spores then passing through the alimentary tract. ----------------------------- Please refer to the
following plates for characteristic structures in the Tuberales: Ascomycota: Euascomycetes: Tuberales Plate 149 = Tuberales ascocarps: Tuber
aestivum, T. rufum & Genea harknessis. Plate 222 = Example Structures --
Discomycetes: Operculatae: Pezizales & Tuberales =
= = = = = = = = = = = = = = |