File:
<basidiomycota.htm> <Index to Mycology> Pooled References <Glossary> Site
Description <Navigate to
Home>
Page 1
True Fungi (Eumycophyta) 1
Basidiomycota (Basidiomycetes, Basidiomycotina)
-- Higher fungi
Heterobasidiomycetes (Contact) Please CLICK on underlined
links & included illustrations for details Use Ctrl/F to search for Subject Matter:
Introduction The Basidiomycota include an immense
variety and number of fungi in the most advanced of all fungal classes. There are several thousand species known
all of which are obligately parasitic on higher plants. They include many
economically important species such as the smuts, rusts, jelly fungi,
mushrooms, puffballs and stinkhorns.
The "shelf fungi"
or bracket fungi also belong
here as well as the less familiar bird's nest fungi. The higher and most familiar members of
this group include the mushrooms, toadstools, puffballs and stinkhorns. The smuts, rusts and jelly fungi
constitute a group that is more primitive than the higher Basidiomycota. A review of the importance of this group
given by Alexopoulos (1952) remains valid to the 21st Century. This class differs from all other
fungi in that they produce their spores, called basidiospores, on the outside
of a specialized, spore-producing body, the basidium. Basidiospores are usually uninucleate and
haploid. Like ascospores they are the
result of plasmogamy, karyogamy and meiosis, the last two of which occur in
the basidium. There are usually four
basidiospores produced on each basidium.
The basidiospores have been considered as homologous to ascospores
because both types develop comparably, and the Basidiomycota have been
regarded as having originated from the Ascomycota. This class of fungi is an
important group of fungi including many harmful species as well as useful
ones. The smuts and rusts are two
groups of parasites causing plant diseases, which destroy a high percentage
of agricultural crops annually. The
most notorious are the stinking smut and black stem rust of wheat. There are many others that attack a
variety of food and ornamental plants.
The higher Basidiomycota are significant in causing diseases of forest
and shade trees and in destroying lumber, railroad ties, etc. In tropical areas where atmospheric
humidity is high it not uncommon to see old automobiles with fruiting bodies
of shelf fungi growing from the skeletons of the cars. Mushroom lovers worldwide seek many Basidiomycota for
food. The cultivation of mushrooms
for food has developed into an industry of considerable proportions and many
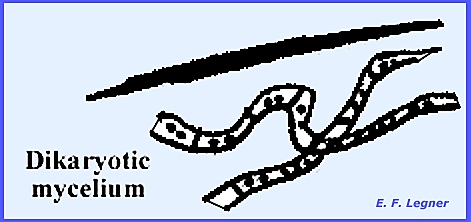
of the wild species are equally good or superior in flavor. Somatic Structures of Basidiomycota Alexopoulos (1952) described the
somatic structures of this class. The
mycelium consists of well-developed septate hyphae, which penetrate into the
substratum and absorb nourishment.
The hyphae individuals are microscopic but they may be plainly seen in
mass. The mycelium is usually white,
bright yellow or orange and often spreads out in a fan-shaped growth. In some forms a number of hyphae lying
parallel to one another are joined together to form thick strands of
mycelium, commonly called shoestrings that technically are known as
rhizomorphs. These strands are
enveloped in a sheath or cortex and behave as a single unit or tissue. The mycelium passes through three
distinct stages of development before the fungus completes its life
cycle. These stages are the primary,
secondary and tertiary mycelia. The primary mycelium
usually develops from the germination of the basidiospore. It may be multinucleate at first, the
nucleus of the basidiophore dividing many times as the germ tube emerges from
the spore and begins to grow. Such a
multinucleate phase of the primary mycelium is short because septa are soon
formed which divide the mycelium into uninucleate cells. The secondary mycelium originates
from the primary mycelium. Its cells
are usually binucleate. The
binucleate condition begins when the protoplasts of two uninucleate cells
fuse, without karyogamy taking place after plasmogamy. The binucleate cell, which is formed,
divides into two daughter cells. The
two nuclei divide conjugately and the sister nuclei separate into the two
daughter cells. An interesting
mechanism that occurs in all major types of Basidiomycota but not in all
species operates to insure that sister nuclei arising from conjugate division
of the dikaryon become separated in the two daughter cells. This mechanism functions through special
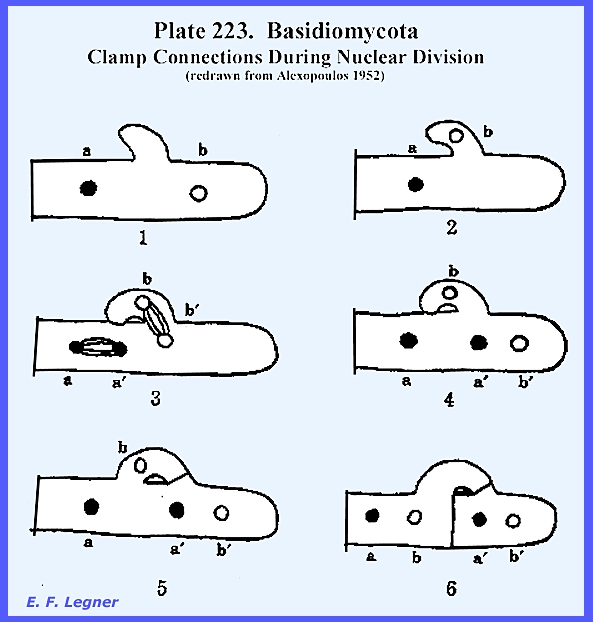
structures called clamp connections, which are formed during nuclear
division. When a binucleate cell is
ready to divide a short branch, the clamp connection, arises between the two
nuclei a and b and begins to form a hook.
The nuclei now divide simultaneously.
The spindle of one division becomes oriented obliquely so that one
daughter nucleus b forms in the clamp connection and the other b' of the
first spindle, which has formed, near the other end of the cell. Meanwhile, the clamp has bent over and its
free end has connected with the cell, so that the clamp forms a bridge
through which one of the daughter nuclei b passes to the other end of the
cell and approaches one of the daughter nuclei (a of the other spindle. A septum forms to close the clamp at the
point of its origin and another septum forms vertically under the bridge to
divide the parent cell into two daughter cells with nuclei a and b in one
daughter cell and nuclei a' and
b' in the other as shown in
the following Plate 223:
---------------------------------- General
Characteristics of Basidiomycota Of some 26,000 species the
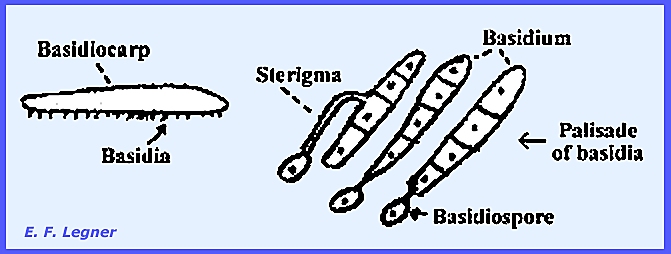
Basidiomycota includes many plant parasites and common fleshy fungi. Their name comes from the "basidium." This structure occupies a place in the
cycle of development and is homologous to the ascus in the Ascomycota. The basidium terminates a dikaryophase and
is the site of nuclear fusion and the seat of meiosis. Basidiospores are primarily uninucleate,
unicellular structures. The basidium
is typically club-shaped and bears spores on sterigmata, which are located on
the outside of the basidium. The
number of spores produced is usually four (in the Ascomycota there are
eight).
Two sub-classes are Heterobasidiomycetes,
which includes a large number of parasitic forms and the rusts; and Homobasidiomycetes,
which more the most conspicuous and commonly known, with many edible
varieties. ---------------------------------- Please refer to the following plates for characteristic
structures in the Basidiomycota: Plate 160 = Basidiomycota:
Hymenium. Plate 161 = Basidium developmental stages. Plate 223 =
Basidiomycota: Clamp Connections
During Nuclear Division ---------------------------------- Five orders of Heterobasidiomycetes are treated here, which
are Uredinales, Auriculariales, Tremellales
and Dacrymycetales
(= frequently included in other orders).
The number of known species as of the year 2010 is only about
520. Many of these are tropical and
the majority is saprophytic. Fruiting
bodies (basidiocarps) are produced in these orders, and in many forms are of
a gelatinous texture that can dry down to a cartilaginous or horny consistency. While the basidia are in all cases mainly
different from those encountered in the Autobasidiomycetes (Eubasidii), they
are not uniform throughout the group.
Actually it is principally on the basis of fundamental differences in
basidial structure that the orders are separated. The Auriculariales, especially through Uredinella and Septobasidium,
are more related to the rusts. The
character of the basidium in the Dacryomycetales allies these forms to the
Agaricales into which order some specialists believe that they should be
incorporated.
The order Uredinales contains
two families: Pucciniaceae
and Melampsoraceae. A typically long-cycled rust produces Five distinct
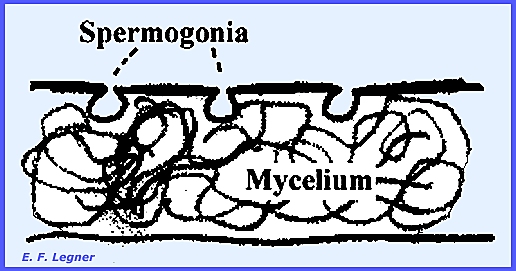
stages in its life cycle in a regular sequence as follows: Stage O = spermogonia bearing spermatia
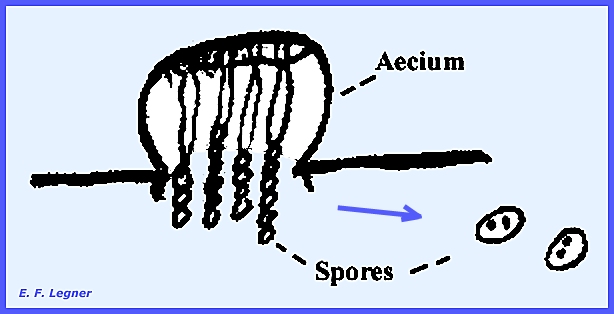
and receptive hyphae. Stage I = aecia bearing
aeciospores. Stage II = uredia bearing urediospores. Stage
III = telia bearing teliospores.
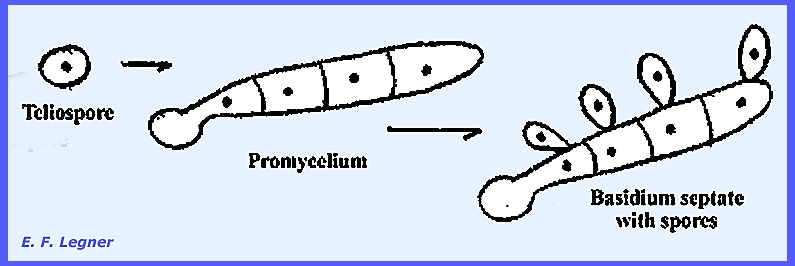
Stage IV = promycelia
bearing basidiospores. ---------------------------------- The family Pucciniaceae generally has
teliospores that are stalked and one- to many-celled. On gymnosperm hosts there is no aecial
stage (Stage I). There are biological
specializations where groups of individuals differ physiologically from other
groups. Subspecies are common and
trinomials are used to distinguish the varieties. In addition there are physiologic races and still further
division of the varieties. Puccinia
gramini-tritici has over 150 physiologic races. The rusts are controlled by eliminating
the secondary host (destruction of barberry in P. graminis) and by the
production of resistant varieties.
But because physiologic races shift from year to year, the breeding
for resistance is a never-ending process.
There are two main types of rusts:
macrocyclic and microcyclid.
The macrocyclic rusts produce one or more types of binucleate spores
in addition to teliospores and they have a long cycle. In the eutype rust all spore types are
present, while in democyclic rusts one spore type is absent. The microcyclic rusts have a short cycle
and they are all autoecious forms.
There are also no spores other than the teliospores (= Stage IV type). The Puccinaceae
is well represented by Puccinia
graminis, which causes "Stem
Rust of Wheat." There are hundreds of millions of bushels
of wheat lost annually in North America due to this pest. The rust is heterocious, heterothallic,
long-cycled or macrocyclic and of the eutype. The heteroecious habit requires the fungus to spend one portion
of its development on another host (autoecious forms complete the life cycle
on the same host). It produces its
aecia on the leaves of members of the Berberidaceae and its uredia and telia
on grasses. The species os composed
of several varieties and numerous physiologic races. European barberry is the dicot
host, which produces spermatia and aeciospores. The mycelium is monokaryotic and penetrates the host, producing
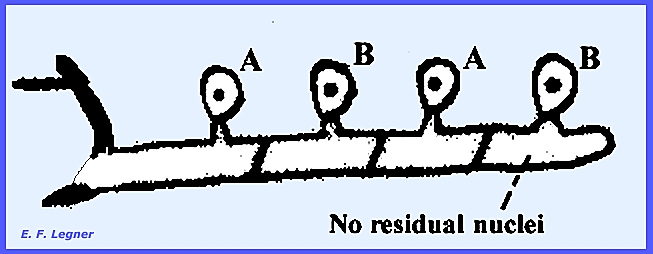
haustoria. There are "A"
and "B" vegetative types of mycelium.
Spermogonia form on the upper
surface of the host.
The spermatia ooze out in a sticky
fluid that is attractive to insect vectors.
At the same time on the lower
surface there are protoaecia.
Another mating type may be
produced on the same leaf or another leaf; and spermatia are blown over to
the sticky fluid of the other type. Flexuous hyphae (receptive
hyphae) protrude from the base of the spermogonium into the sticky mass and
receive the spermatia of the opposite mating type.
The nuclei eventually reach the
protoecium in a process that is not clear; and the protoaecium is then
changed into an aecium. Aeciospores are produced in the aecium,
which is cup-like in shape. Spores
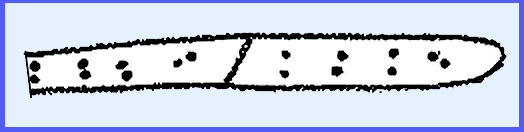
are catenulate and each spore is dikaryotic with one "A" and one
"B" nucleus.
The origin of aeciospores is from a single cell at the base of
the aecium. It marks the beginning of
the dikaryophase. Nuclei move into
extensions in pairs, which is very similar to that found in ascogenous
hyphae. The Monocot Host of
Puccinaceae After indirect penetration of the germ tube of the
aeciospore through the stoma of the monocot host (= wheat & other
grasses), the mycelium spreads. At
this time it is dikaryotic. It should
be noticed that this is in contrast to the situation on a dicot host where
penetration of the germ tube is direct and the mycelium is monokaryotic.
A pustule called a uredium
is formed which
produces urediospores. They are 2-nucleate (dikaryotic).
Urediospores
function like conidia and constitute the vegetative stage of the rust. On the same mycelium are formed the teliospores,
which are produced in the uredium that is now called a telium. The teliospore is at first dikaryotic but
nuclear fusion occurs forming a diploid.
Colors of the respective spores are teliospore = brown or black and
urediospore = orange.
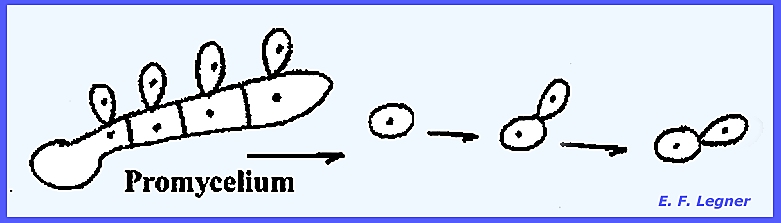
A germ tube
protrudes from each cell of the teliospore,
which remains attached to the telium.
Each cell of the tube hs but one nucleus.
Strigmata
are produced, which bear basidiospores.
Basidiospores are uninucleate and either "A" or
"B" (there are two of each type).
Puccinia graminis is a eutype in that
all spore forms are represented in the life cycle. This species is also heteroecius, heterothallic and
macrocyclic-eutype. ---------------------------------- In other Puccinia species
the teliospores are similar throughout.
Most have a well-developed cupulate aecium and the life cycles differ
among species. For example, Puccinia
podophylli, causing Mayapple Rust, is a democyclic
rust. There is no uredial stage
(Stage II), and it is an autoecious form (both mono- and dikaryotic mycelium
on the same host). Puccinia malvacearum,
causing Hollyhock
Rust is microcyclic. Uromyces spp. have
one-celled teliospores.
Phragmidium sp. has a uniquely-shaped spore
Gymnosporangium juniperi-virginianae
causes "Cedar-Apple Rust,"
and G. globosum causes "Hawthorn Rust. "Both of these rusts re demicyclic
and heteroecious. Red cedar serves as
the alternate host for both species and conspicuous galls form on it as a
response to invasion by either parasite.
The telial stage develops on these galls in the spring. In life these spore horns are bright
orange. ---------------------------------- The family Melampsoraceae has sessile teliospores
and is heteroecious (conifers are the alternate host). The following genera will reveal the characteristics
of this family: Uredinopsis spp. are the
"Fern Rusts." Gymnosperms, Abies spp., are the
aecial host in North America (Stages
0 & I); Fern is the uredial host.
This is a macrocyclic group. Melampsora spp. have teliospores
formed in crusts; they are one-celled and stalkless.
Urediospores have capitate
paraphyses arranged with them in the uredium. The gynnosperm host is usually larch for the aecial stage. This is a macrocyclic, eutype group. Coleosporium spp. occur on
aster in the uredo stage and on pine needles in the aecial stage. There is a peridermoid aecium
(blister-like). They occur on pines
that have two needles. Urediospores
form chains. There is no external
promycelium produced from the teliospore.
Instead karyogamy and meiosis occur in the teliospore. This is a macrocyclic eutype group. Coleosporium solidaginis is a
common variety in North America. It
is heteroecious, producing tongue-like orange-colored aecia on the leaves of
2-needled pines. Its uredial and
telial stages occur on gondenrod, asters, etc. Cronartium spp.: C. ribicola incites White Pine
Blister Rust. They are
found on pines with more than two needles.
They are long-cycled, heteroecious and of the eutype. Telial and uredial stages occur on Ribes
and Grossularia (currant & gooseberry); aecial stages are on white
pine. C. ribicola is common on
white pine (Pinus strobus).
The teliospores are laid side-by-side and end-to-end. C. quercuum occurs on oak where
there are the uredal and telial stages.
The teliospores are like C. ribicola.
Cronartium
quercuum is
another common species in North America.
This rust stimulates the production of large aecial galls on jack pine
branches. The uredial and telial
stages occur on various species of oak.
---------------------------------- Please refer to the following plates for
characteristic structures and Life Cycles in the Uredinales: Basidiomycota:
Heterobasidiomycetes: Uredinales Plate 168 = Basidiospores: Uredinella coecidiophaga Plate
169 = Rust spermogonia (2 types) Plate
171 = Uredia: Two types Plate
172 = Teleutospores: Uromyces, Pileolaria, Puccinia,
Uropyxis, Xenodochus, Phragnidium, Nyssopsora, Ravenelia. Plate
173 = Life Cycle -- Puccinia graminis Plate
174 = Telium of Cronartium ribicola Plate 224 =
Diagnostic Characters-1 -- Uredinales:
Pucciniaceae Plate 225 =
Diagnostic Characters-1 -- Uredinales:
Melampsoraceae Plate 226 =
Diagnostic Characters-2 -- Uredinales:
Pucciniaceae Plate 227 =
Diagnostic Characters-3 -- Uredinales:
Pucciniaceae Plate 228 = Diagnostic
Characters-2-- Uredinales:
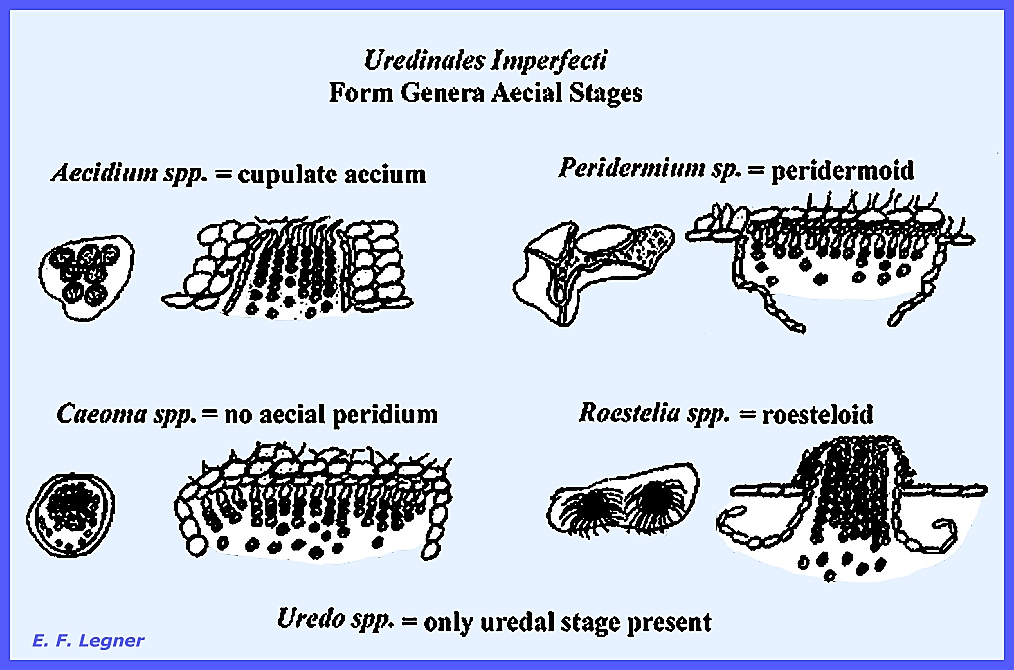
Melampsoraceae ---------------------------------- The Uredinales Imperfecti is a group that
contains the Imperfect
Rusts. Because rusts have
a polymorphic life cycle their classification is complicated. As found in the Ascomycota, but to a
greater degree, imperfect stages of most rusts are more likely to be found
than perfect stages in certain seasons.
As the classification of the rusts is based on teleutospores, it is
difficult to such structures if they are not known or not available (teleutospores are
thick walled resting spores in which karyogamy occurs; they are part of the
basidial apparatus). Also because in
heteroecious rusts the teleutospores occur on a different host from that on
which the aeciospores occur complicates the situation even more. Therefore, this has led to the creation of
an artificial group, the Uredinales Imperfecti in which the aecial and
uredial stages are temporarily classified until the teleutospores are
discovered.. There is no perfect
stage known (= telial stage). Various
genera show the characteristics of these fungi in the following Fig. 385:
---------------------------------- Please refer to the following plates for characteristic
structures and Life Cycles in the Uredinales Imperfecti Basidiomycota:
Heterobasidiomycetes: Uredinales Imperfecti Plate 170 = Aecia types: Caeoma, Aecidium, Roestelia
& Peridermium. Plate
177 = Life Cycle -- Tilletia caries. ---------------------------------- The order Ustilaginales
includes over 1,200 species in over 35
genera. These are the "smuts,"
which are important plant pathogens on cereal crops and vegetables. None are obligately parasitic and there is
an intercellular mycelium with or without haustoria. The species have a simpler life cycle than
rusts and the dikaryophase is terminated by the teliospore stage. The former order Uredinales differs by
having over 6,000 species none of which are obligately parasitic; and
teliospore production occurs with one or more probasidia, which bear
basidiospores (sporidia). Unlike the
rusts smut fungi can be cultivated on artificial media. They exhibit no trace of sex organs. Among the Heterobasidiomycetes only in smuts
and rusts is no basidiocarp produced. Teliospores in the Ustilaginales
are conspicuous, dark masses (= pustules or spore masses). The usually black
spores are often formed in the ovary of the host and produced in the
millions. Hypertrophy is commonly
induced in the host, but sometimes it is absent. In one very serious disease, "corn
smut", hypertrophy occurs throughout the
plant. Some smuts sporulate only in
the ovaries of the host, but others form spores in other parts of the plant. The smut spore is commonly spherical. Two nuclei fuse prior to formation of the
basidium and spores are often held together in clusters, or spore balls, of
they may be free and separated.
Spores are retained in the host until the rupture of host tissue, and
they have been given various names such as "winter spores", "smut spores",
"brand
spores", "teliospores" or "chlamydospores." In the smuts conjugation may occur
between compatible basidiospores. The
one nucleus migrates into the other basidiospore. A basidiospore may germinate sending a hypha into the
host. Growth of this hypha is
weak. But if the mycelium of one basidiospore
comes into contact with the mycelium of another spore, they will join
together and by hyphal anastomosis set-up the dikaryophase. Some basidiospores may conjugate right on
the basidium and 2N hyphae are produced right on the basidium (= "H"
connections). The mycelium may
have multinucleate cells within, although the nuclei always occur in pairs. In destroying host tissue the
hyphae may be very dense and profuse, and branching may occur. Eventually the hyphal cells disarticulate
in defined areas with subsequent rounding up and formation of binucleate
spores. Some smuts attack only the
seedling stage of a host, which becomes resistant with age. Some of the smuts on cereals are like
this. The corn smut fungus infects
the older leaves of maize and also the tassel and stem. Attack occurs on new tissue only, however. The teliospore lands on a style
and then grows down into the host ovary and fight on through the ovary
wall. Systemic infections occur in
many forms. Onion smut attacks the
growing points of the seedling. Some
smuts enter the embryos of their hosts and then remain dormant until planting
time. The maize kernel infected with
corn smut may be expanded to 100-times its normal size. On smut fungus remains in the soil. Bunt, oat smut or loose smut of barley all
have a terrific production of spores.
Surface sterilization of seed or grain with Formalin can control these
fungi. Loose smuts that attack deep
within the host tissue require hot water treatment to be controlled. Developmental Cycle
of Ustilaginales The mycelium is typically dikaryotic
in the host, but often there may be multinucleate cells that occur in pairs.
They are predominantly heterothallic.
A tetrapolar compatibility is
present: AB, Ab, aB, ab. However,
only the AaBb form is compatible. Teliospores arise when dense knots
of hyphae form, each cell of which becomes definitely binucleate. The walls disappear and free the protoplasts,
which round up and develop a new wall by themselves. These develop into the teliospores. Teliospores, which are diploid, form the
overwintering stage. Families in the Ustilaginales
are separated on the basis of the form of the promycelium and the way in
which the basidiospores (sporidia) are borne on this structure. Genera are defined primarily on teliospore
characters, including such features as whether the spores occur singly, in
pairs, or in spore balls. Two
families presented here are Ustilaginaceae and
Tilletiaceae. The principal differences are shown in Fig. 387 and Fig 388 as follows:
Genera are defined primarily on
teliospore characters, including such features as whether the spores occur
singly, in pairs, or in spore balls. Ustilaginaceae
Tilletiaceae
The number of basidiospores in the
Tilletiaceae is indefinite because mitosis occurs leaving a nucleus behind in
the promycelium and one in the basidiospore. A few species of smuts produce
conidia even though the principal reproductive struture is still the
teliospore. The conidia are dikaryotic
and give rise to new mycelia. In the Ustilaginaceae, Ustilago
hordei causes "Covered Smut of Barley and
Oats." Here
basidiospores bud out from cells of the septate promycelium.
In the Tilletiaceae, Urocystis
cepulae causes "Onion Smut." This fungus will not germinate at high
temperatures, hence in warmer climatic areas the planting of onions is timed
in accordance with the least prevalence of the disease. The spores occur in the soil so that a
sterilizing solution is dribbled in the seed row at planting time, which
remains effective through the several weeks when the onion is susceptible. For controlling smut fungi in
general it is important to understand their habits. In Tilletia caries, Ustilago hordei and U.
avenae the spores occur on the surface of the grain. As the host is susceptible only at the
very young stage, surface sterilization with Formalin kills the spores. When the fungus occurs in the embryo,
surface sterilization will not kill the spores, but rather hot water
treatment is effective. When the
mycelium of a smut is localized in development in the meristematic regions,
resistant varieties are the best means of control. ---------------------------------- Please refer to the following plates for characteristic
structures and Life Cycles in the Ustilaginales: Basidiomycota:
Heterobasidiomycetes: Ustilaginales Plate 175 = Teleutospores: Ustilago levis, U. maydis, Tilletia
caries, Urocystis cepulae, Tuburcinia trientalis, Thecaphora seminis-convolvuli & Schroeteria
delastrina. Plate
176 = Life Cycle -- Ustilago maydis. Plate
229 = Diagnostic Characters:
Ustilaginales: Ustilaginaceae Plate 230 =
Diagnostic Characters:
Ustilaginales: Tilletiaceae ---------------------------------- The order Auriculariales
is distinguished by
having a transversely septate basidium that is comparable to the ascocarp and
closely resembles that found in the Uredinales. Auricularia (Hirneola), a saprophyte on stumps,
dead tree trunks, etc., is one of the commonest and best-known
representatives of the order. Pilacre
is also a frequently encountered saprophyte.
Eocronartium musciola, on moss, is one of the few well-known
Bryophyte parasites. Septobasidium
parasitizes scale insects. Families
of the Auriculariales that have been recognized are Auriculariaceae and
Septobasidiaceae. There is a dikaryophase
established through hyphal anastomosis.
The basidiocarp bears the dikaryophase and also terminates it in the
basidiospores. Basidiospores
germinate to produce a monokaryotic mycelium. Hyphal anastomosis then sets up the dikaryophase. The absorptive dikaryotic mycelium here is
quite different than that found in the Ascomycota: Taphrinales. There are
no sexual organs in all the Basidiomycota generally, and only the Uredinales
have spermatia. ---------------------------------- The family Auriculariaceae has fruiting bodies that
vary from a simple weft of hyphae to well-developed, large fruiting
bodies. Most are saprobic with a few
being parasitic on mosses and the roots of flowering plants. The Genus Auricularia represents this group, which is
most closely related to the rusts and smuts.
Ear-like fruiting bodies with a gelatinous texture are produced on the
outside of wood. Sterigmata of
variable length are formed, and the basidiospores that they bear are all
situated on a similar plane.
The Genus Pilacre has small,
mushroom-like basidiocarps, but these are not gelatinous. The Genus Eocronartium attacks the gametophyte generation
of mosses. These fungi resemble Cronartium telia on the
gametophyte. ---------------------------------- The family Septobasidiaceae includes
about 165 species of which the genera Septobasidium and Uredinella re
parasitic on scale insects. Members
of the family resemble the Auriculariaceae in that they produce transversely
septate basidia. They differ from
most Auriculariaceae in that the fruiting bodies of the Septobasidiaceae are
not gelatinous and in some parts of their biology. The Auriculariaceae are saprobic or parasitic on plants whereas
the Septobasidiaceae are parasitic on insects. Of evolutionary significance is that in most species of Septobasidium
the wall of the hypobasidium is very thick and the whole structure resembles
a spore. The hypobasidium eventually
emits an elongated epibasidium that soon becomes transversely septate into
four cells. Each of these cells then
produces a sterigma that in turn forms a basidiospore. These hypobasidia appear similar to the
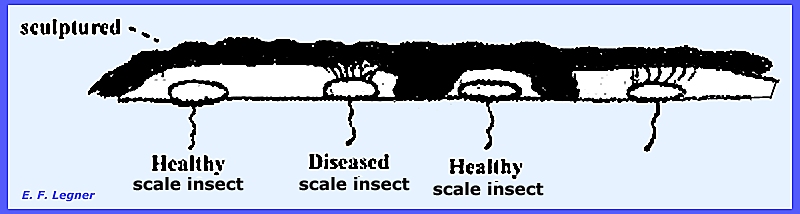
teleutospores of the Uredinales. The Genus Septobasidium
attacks whole colonies of scale insects.
The basidiocarp is distinctively sculptured and covers the entire
colony of insects. It will not attack
healthy insects and infection occurs only via spores.
When the healthy scale insects
mature they crawl out of their "cage" through a channel in the
basidiocarp. Sporulation is timed
with the emergence of the crawlers, which acquire spores as they rest on the
surface of the basidiocarp. The Genus Uredinella is similar to Septobasidium
except that only one insect at a time is attacked. Basidiospores are formed from a probasidium in both species and
the attacked scale insects are not severely harmed. ---------------------------------- Please refer to the following plates for characteristic
structures and Life Cycles in the Auriculariales: Basidiomycota: Heterobasidiomycetes:
Auriculariales Plate 166 = Life
Cycle -- Auricularia auricula. Plate
167 = Structures of Septobasidium
fumigatum. Plate 231 = Example
Structures -- Auriculariales, Tremellales, Dacryomycetales ---------------------------------- The order Tremellales includes the "Jelly
Fungi." Their basidiocarps vary from crust-like to
stalked, and in some forms the basidiocarp is only a thin layer of gelatinous
hyphae that produce the basidia. One
family Tremellaceae has been
recognized. The life cycle is similar
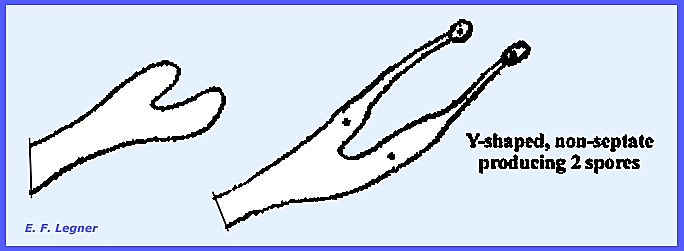
to the Auriculariales except that the basidium differs. Vertical septations occur in the basidium
and serigmata grow out and produce basidiospores at their apex.
---------------------------------- Please refer to the following plates for characteristic
structures and Life Cycles in the Tremellales Basidiomycota:
Heterobasidiomycetes: Tremellales: Plate
165 = Life Cycle -- Exidia spiculosa. Plate 231 = Example
Structures -- Auriculariales, Tremellales, Dacryomycetales ---------------------------------- The order Dacryomycetales
are also Jelly Fungi
whose fruiting bodies are easily recognized as small bodies, bright yellow or
orange jelly-like or waxy. They occur
on the branches or trunks of living or dead trees. There is a strong tendency to produce secondary basidiospores
in this order, which may continue growing in a yeast-like phase if cultured. A single family the Dacryomycetaceae
has been recognized. ----------------------------------
Please
refer to the following plates for characteristic structures and Life Cycles in
the Dacrymycetales: Basidiomycota: Heterobasidiomycetes:
Dacrymycetales Plate 162 = Basidiospore formation: Calocera cornea. Plate
164 = Life Cycle -- Dacrymyces
deliquescens. Plate 231 = Example
Structures -- Auriculariales, Tremellales, Dacryomycetales ================ |















{kind=link}
{kind=link}