File:
<zygomycota.htm> <Index to Mycology> Pooled References <Glossary> Site
Description <Navigate to
Home>
Page 1
True Fungi (Eumycophyta) 1
Zygomycota
(Phycomycetes) -- Zygote fungi
(Contact) Please CLICK on underlined
links & included illustrations for details Use Ctrl/F to search
for Subject Matter: Sample Examinations
The Zygomycota includes a group of
organisms that show a tremendous variation in structure. There is some question as to whether these
are advanced organisms that have degenerated, or whether they are truly
primitive. Subclasses are
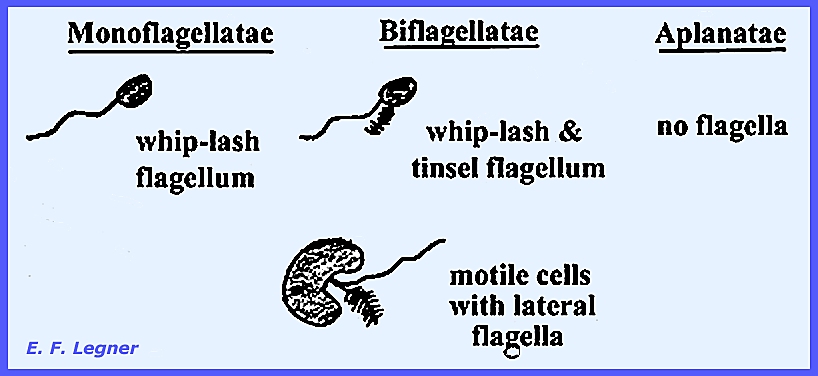
Monoflagellatae, Biflagellatae and Aflagellatae. It is suspected that these three groups might have evolved
independently. Monoflagellatae The order Chytridiales has been considered at
the evolutionary bottom of the Monoflagellatae and are thus among the lowest
of the True Fungi. There are over 40
genera and several hundred described species, all distributed among over ten
different families. The majority of
species are aquatic, but some species are common in some soils and may also
be found in other terrestrial habitats.
A considerable number are known to live as saprophytes on submerged
organic debris; others parasitize algae, especially freshwater algae or
aquatic filamentous fungi. Some
species grow on microscopic animals, or on pollen grains that have fallen
into the water. Finally some attack
terrestrial angiosperms, and among these there are incitants of some
important diseases of food plants. Among the
simplest members of the Chytridiales the body is a single globular cell, and
the entire structures is eventually used up in the formation of reproductive
bodies. Such organisms are often
called holocarpic. Among the more advanced species the
thallus is polycentric, a number of nucleated centers being connected by a
system of generally enucleate rhizomycelial threads. The nucleated areas are transformed into
or serve as points of origin for reproductive organs. The Genus Olpidium parasitizes algae,
fungi, pollen, small aquatic animals and some higher plants. The life cycle has been worked out on
cabbage, where the organism is unicellular, uninucleate and
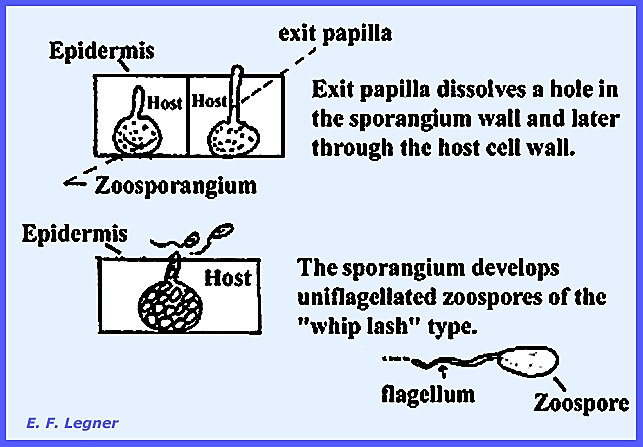
intracellular. An exit
papilla dissolves a hole in the sporangium wall and later through the host
cell wall. The sporangium develops uniflagellated zoospores of the “whip
lash” type. The organism is
Holocarpic where the entire vegetative thallus is used up in reproduction (Eucarpic =
only a portion of the thallus is used in reproduction).
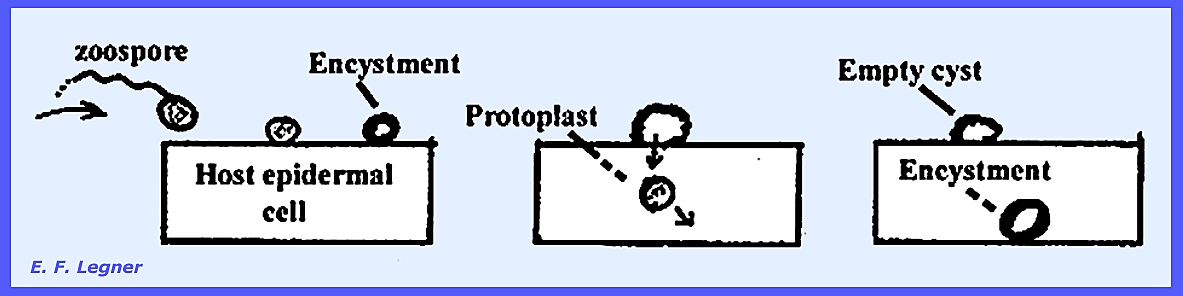
The zoospore
(or planospore, swarm spore) lands on another cabbage root epidermal cell and
loses its flagellum and builds a wall around itself. The wall remains on the epidermis and the protoplast
is discharged into the cell. It may
go in for a few layers of cells before building up another wall around
itself. Germination follows.
In some cases
the zoospores may act as facultative gametes. Two gametes may fuse to form a zygote (diploid). Rather than the protoplast penetrating the
host in this case only, the entire zygote will enter, no portion being left
outside. A wall is secreted around
the zygote and the overwintering stage is formed. In the spring meiotic division occurs and produces a
multinucleate structure (haploid nuclei).
The zygote then turns loose many uniflagellated zoospores. When gametes are produced, the
“sporangium” may best be referred to as a gametangium). [See PLATE 15 for Life Cycle of Olpidium viciae]. The Genus
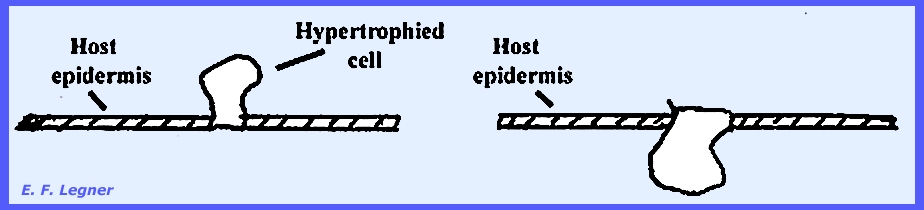
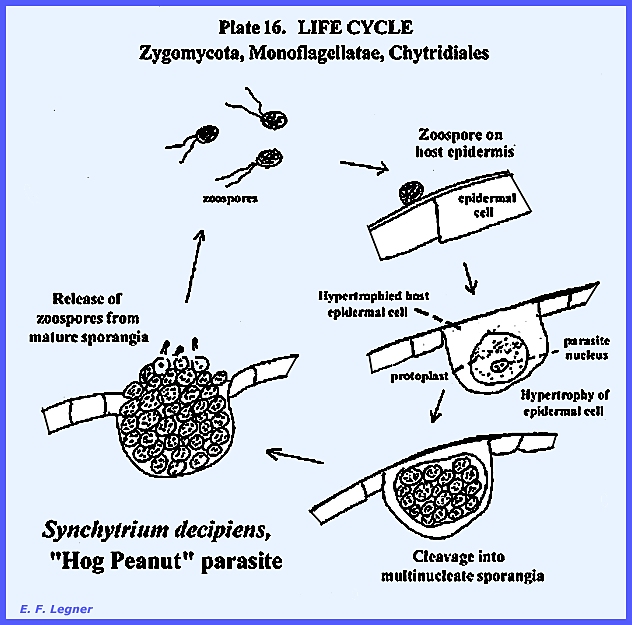
Synchytrium has species that parasitize higher plants. In the reproductive phase there is a
cluster of sporangia involved. The
parasite induces hypertrophy of the epidermal cells of the “hog peanut” host.
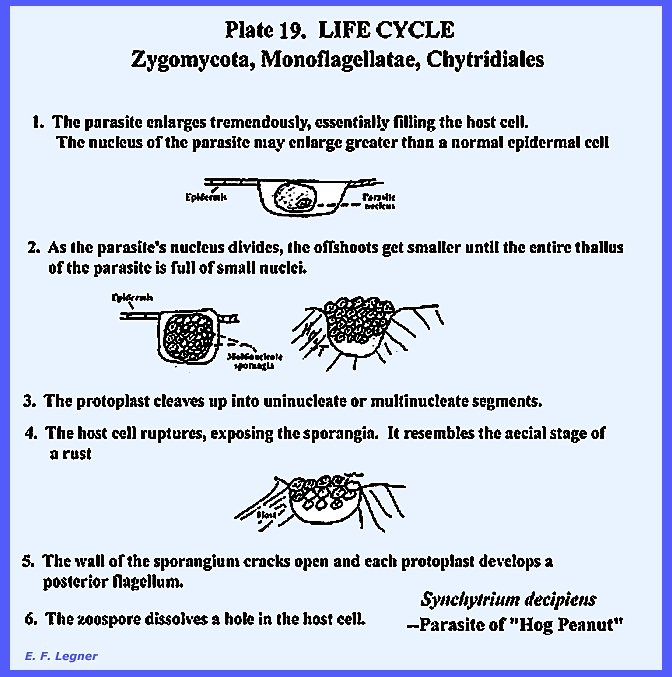
The life cycle of Synchytrium
decipiens, is shown as follows in PLATE 16 & PLATE 19:
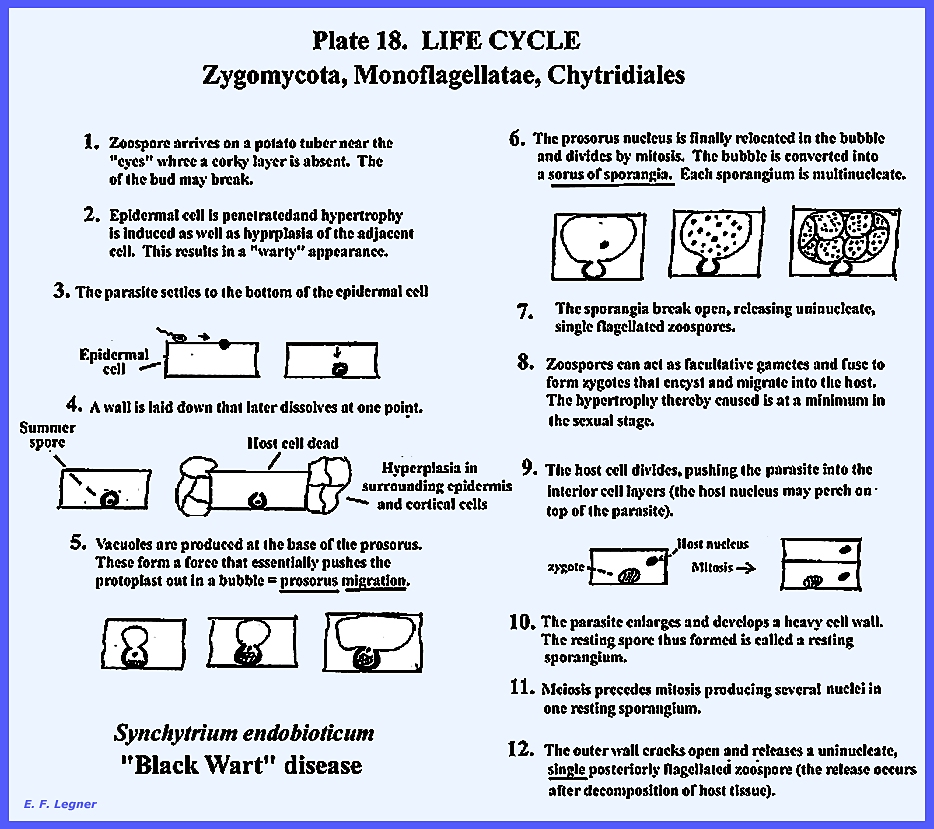
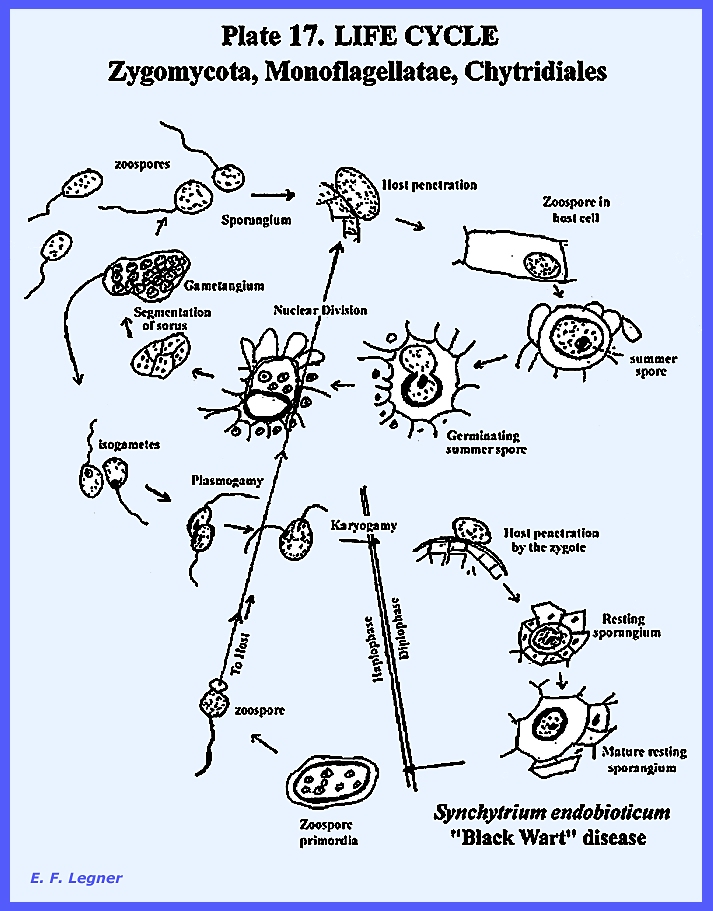
The life cycle of Synchytrium endobioticum is shown in PLATE 17 and PLATE
18:
Another species, Synchytrium
cellulare, parasitizes false wood nettle leaves. It is similar to S. endobioticum in
having a sexual and asexual stage, but there are 9-16 sporangia in a sorus (S.
endobioticum has 7-9). When septa occur in the
Zygomycota, which may happen in an older mycelium, they are complete and
without a septal pore. The
flagellation of motile cells is the basis of the Subclass classification:
The vegetative thallus of
Chytridiales is single-celled (holocarpic).
There is a cell wall, typical of a true fungus, and single-flagellated
motile cells. Synchytrium is one step
ahead of Olpidium in that it possesses a character where each thallus
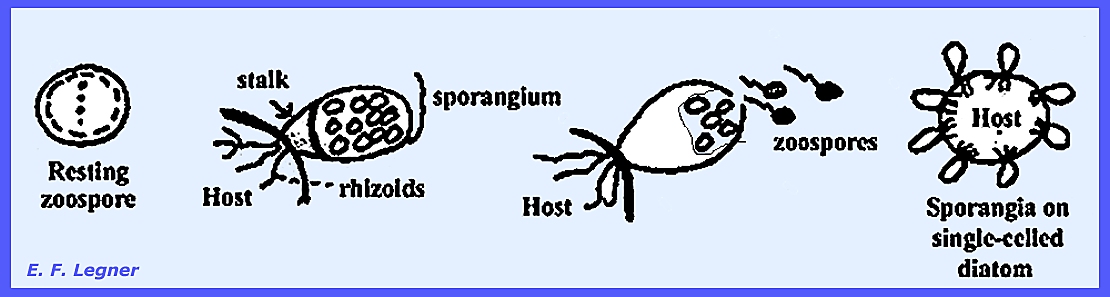
goes into several sporangia. Podochytrium cornutum parasitizes diatoms of the
Genus Stephanodiscus. There is
a single uniflagellated zoospore that comes to rest on Stephanodiscus. It then sends out rhizoids, which
penetrate diatoms, presumably through the pores in the walls. Rhizoids may be called haustoria, but they
are not “hyphae” because there is no nucleus present. More than one parasite (zoospore) may
infect one diatom cell at any given time.
P. cornutum is a monocentric and eucarpic organism.
The Asexual Stage in the Genus Rhizophidium
has zoospores with a single posterior flagellum that land on a host cell (an
alga).
The spore sends out rhizoids and
the protoplasm divides into more nuclei and then cleaves into uninucleate
divisions. These then become
zoospores. They are eucarpic. The Sexual stage of Rhizophidium
has two zoospores, which land side by side on an alga. One will enlarge and the wall between the
two cells will rupture. The
protoplasm of the small one erupts into the larger one. A resting sporangium with a hard
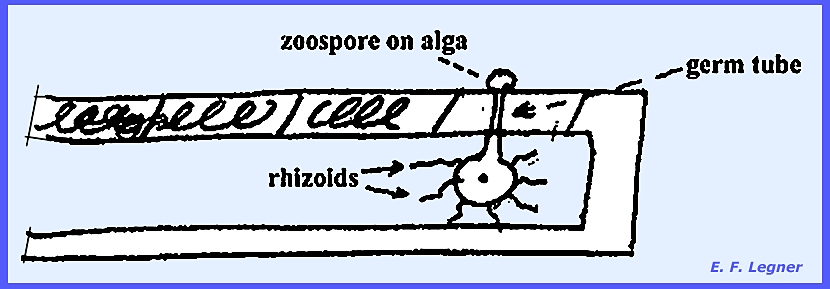
surrounding wall is formed and the zoospores are released thereafter. The Genus Entophlyctis has motile
zoospores that land on algal filaments and send germ tubes through the host
cell wall.
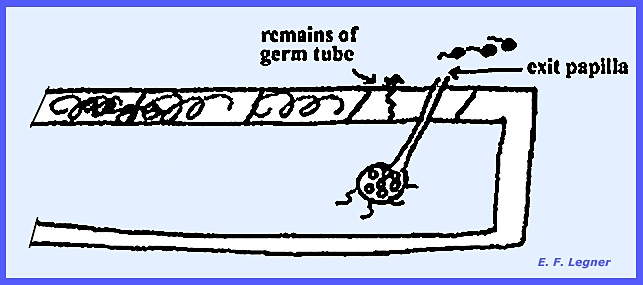
Nuclear division precedes
cleavage. The swollen section is
abscised from the germ tube, and an exit papilla is sent up to the surface
where zoospores are released. There is no sexual stage and the organism is Eucarpic and Monocentric).
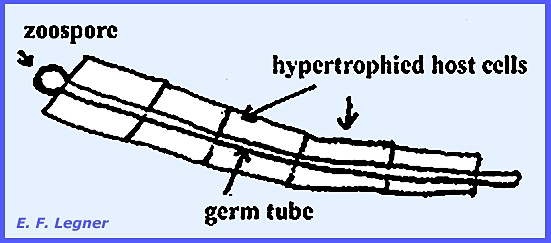
Urophlyctis alfalfae causes “Crown
Wart” disease. It differs from Physoderma zeae-maydis by
inducing hypertrophy. A single
zoospore alights on a host epidermal cell and sends out a germ tube, causing
the hypertrophy. The tube may extend
to more than one cell (= Polyphagous).
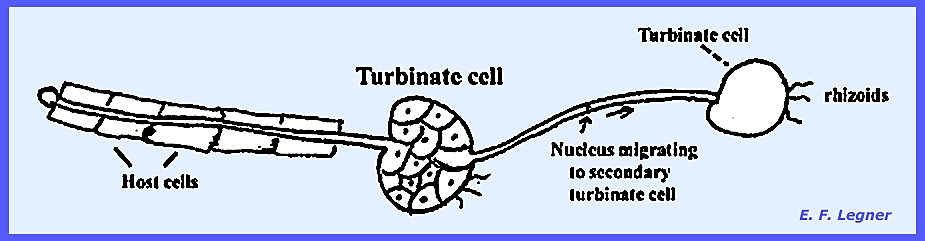
The nucleus of the zoospore
migrates to a turbinate
cell where it will divide into 5-8 nuclei, which cleave into as
many cells. Each of the cells of a turbinate
cell may send out a network of germ tubes with their respective turbinate
cells. The organism is polycentric. The turbinate cells mature and a
system of sparsely branched rhizoids develop at the end of each turbinate
cell. Eventually one of the cells at
the end of a turbinate, instead of giving rise to a rhizo-mycelial system,
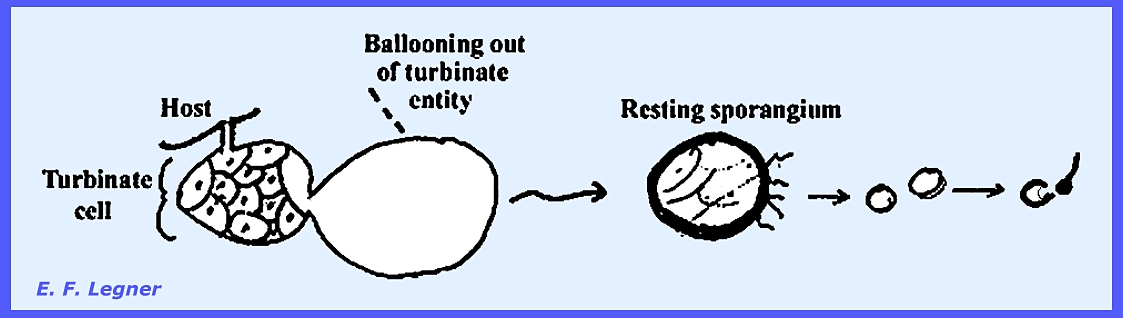
balloons out to form a structure larger than the turbinate cell itself.
The nucleus divides; a thick wall
is laid down and forms a resting sporangium.
A halo of rhizoids is also present.
Often the host tissue may wall-off a cluster of resting sporangia with
corky tissue. Eventually the host
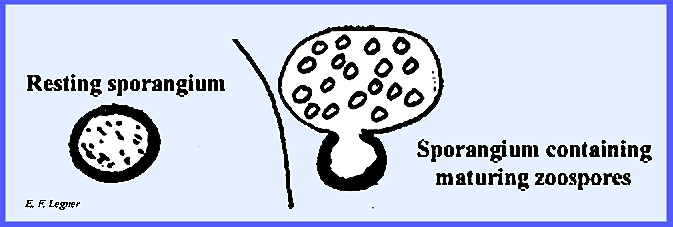
disintegrates, releasing the resting sporangia. These will split up into many multinucleate sporangia. Each then cleaves into uninucleate
portions that give rise to a zoospore.
The zoospore may land on a host’s epidermal cell where it sends out
some rhizoids. This structure then
forms a sporangium (ephemeral sporangium), which in turn releases zoospores that
begin the vegetative mycelium over again.
There is no evidence of a sexual stage.
Physoderma zeae-maydis causes “Brown
Spot” disease of
maize. The sporangium is
slipper-shaped. As soon as the
sporangium becomes empty an internal proliferation occurs and a new sporangium
develops from a remaining nucleus of the old sporangium. The new sporangium releases
zoospores. This process may be
repeated several times. The
slipper-shaped structure may be considered a gametangium. As the germ tube goes through a
maize leaf, there is no hypertrophy.
The resting sporangium migrates into the upper sorus portion and
cleavage occurs forming the sporangium.
Each entity gives rise to one zoospore.
------------------------------------------- Please see
following plates for Life Cycles and Structural characteristics in the Chytridiales: Zygomycota: Monoflagellatae:
Chytridiales Plate
15 = Life Cycle -- Olpidium viciae Plate
16 = Life Cycle -- Synchytrium decipiens #1 Plate
19 = Life Cycle -- Synchytrium decipiens #2 Plate
17 = Life Cycle -- Synchytrium endobioticum Plate
18 = Life Cycle -- Synchytrium endobioticum #2 Plate
20 = Life Cycle Podochytrium
cornutum Plate
21 = Life Cycle -- Chytridiales: Endophlyctis Plate
22 = Life Cycle -- Chytridiales: Rhizophidium
couchii Plate 78 = Life Cycle2 -- Rhizophidium couchii. Plate
23 = Life Cycle -- Chytridiales: Urophlyctis
alfalfae Plate
24 = Example Structures -- Chytridiales:
Sexual Reproduction Plate
25 = Example Structures -- Chytridiales:
Synchytrium spp. Plate
26 = Example Structures -- Chytridiales:
Physoderma, Podochytrium, Urophlyctis. Plate 74 = Thallus of Chytriomyces
aureus. Plate 75 = Flagella of Zygomycota: Whip lash, Tinsel and combination. Plate 76 = Life Cycle -- Olpidium viciae. Plate 77 = Life Cycle -- Synchytrium endobioticum. ================================= The order Blastocladiales
has an arbusculate mycelium. Species are usually aquatic, but some may
occur in moist soil. The zygotes do
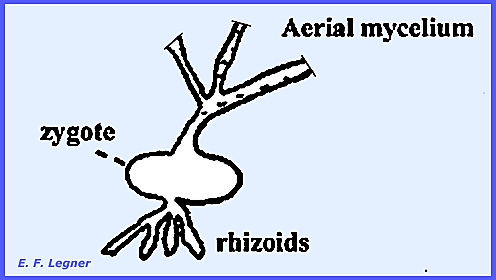
not have a hard wall. Allomyces arbusculus is a typical representative of
this order. It lives in fresh water
or moist soil and the thallus is visible with the naked eye. The base of the mycelium is attached to a
plant thallus by rhizoids. It has a
coenocytic mycelium. During the life cycle the trunk
will branch and the tips of branches differentiate into hairs of sex
organs. The bottom differentiation is
reddish and small (male) while the top one is clear and larger (female).
The thallus is multinucleate and
has N chromosomes (gametophyte generation on N thallus). Smaller differentiation cleaves into many
protoplasts that form gametes with a single posterior flagellum (male). The larger one cleaves into a smaller
number of larger gametes, which are more sluggish in their movement
(female). Two gametes eventually
fuse, giving a zygote. The zygote is
biflagellated for a short period of time.
A system of rhizoids is sent out from the basal end while aerial
hyphae are sent out from the anterior end.
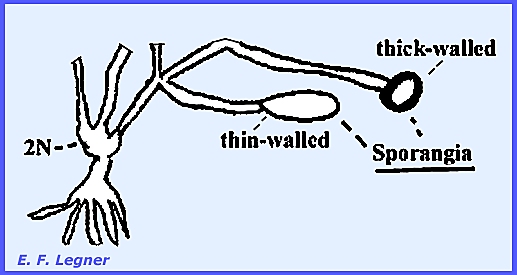
This thallus is 2N.
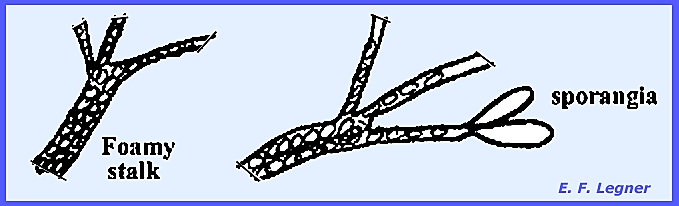
The tips of the 2N thallus can
differentiate into either of two structures.
The first is a thin-walled sporangium that may occur singly or in a
chain. It gives rise to a 2N
zoospore, which in turn gives rise to another 2N thallus.
The second structure is a thick-walled
sporangium that possesses characteristic pits. It can survive for years in the dry state. In a moist environment the multinucleate
protoplast will divide and release uniflagellated motile cells. Meiosis is believed to take place in the
thick-walled sporangium. Spores are
1N and either fuse to form a 2N thallus or they may simply give rise to a 1N
thallus without fusing. The gametes
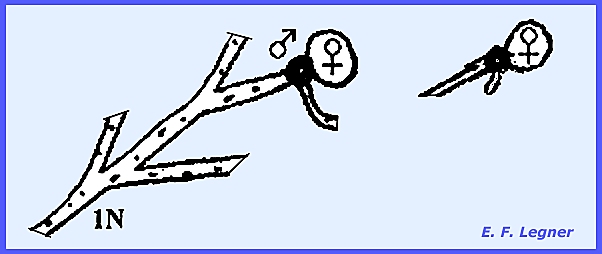
involved are called anisogametes that only differ in size. An Alternation of Generations is
shown by the genus Allomyces.
In the Genus Blastocladia there is an extensive system of
rhizoids. They are an aquatic genus
with an arbusculate mycelium that usually branches dichotomously. During the life cycle the tips of the
mycelium branches giving rise to clavate or elongated, thin-walled sporangia
(think-walled = 2N). Or they may
develop spheroid, thick-walled sporangia, which are also 2N. There is no sexual stage or resting spore
and no Alternation of Generations.
===================================== The order Monoblepharidales has a species that
parasitizes nematodes and another the larvae of mosquitoes. Their zygotes develop a hard wall. Monoblepharis is a
representative species that will develop a coenocytic mycelium with a few
rhizoids at the base and the indication of an arbusculate form. The protoplasm in the stalk becomes highly
evacuolated, which is a good diagnostic character.
One or more sporangia develop at
the tip, and these release zoospores.
There is an internal proliferation where a new sporangium arises from
an old sporangial wall, an event that may reoccur 3-4 times. At the ends of other branches male and
female gametangia will differentiate.
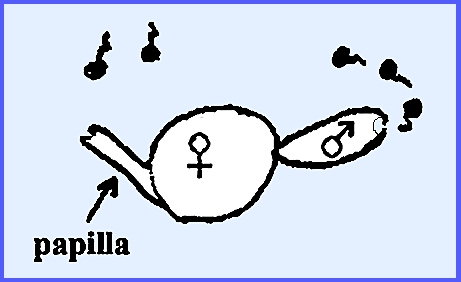
The female gamete is uninucleate
and referred to as an oösphere. At maturity
a papilla will jet out to one side of the oösphere, which allows an entry
point to male gametes. The male
gametangium is multinucleate and cleaves out into many single
posterior-flagellated gametes (antherozoid).
These swim toward the papilla of the female gametangium.
Oögamy is exhibited in this
genus. Karyogamy has not yet
occurred, and the binucleate “zygote-to-be” crawls out to the end of the
papilla and drops off. The nuclei
fuse and then the structure germinates by sending out a germ tube. The mycelium may be either 1N or 2N,
depending on when meiosis occurred.
The 1N mycelium is probably most common. There is no Alternation of Generations.
In another genus, Monoblepharella, the
“zygote-to-be” swims around a bit before forming the resting spore. ---------------------------------------- Please see
following plates for life cycles in the Blastocladiales and Monoblephariales: Zygomycota: Monoflagellatae:
Blastocladiales Plate
27 = Life Cycle – Blastocladiales:
Allomyces arbusculus Plate
28 = Life Cycle -- Blastocladiales:
Blastocladia Plate 79 = Life Cycle -- Allomyces javanicus var. macrogynus. Zygomycota: Monoflagellatae:
Monoblepharidales Plate
29 = Life Cycle -- Monoblephariales:
Monoblepharis Plate 80 = Life Cycle -- Monoblepharis polymorpha. ---------------------------------------- |