Fuel
Use During Hovering Flight | Metabolic Substrate and Oxygen Consumption
|
High Performance Muscle Power Output
Metabolic Substrate and Oxygen Consumption
Research conducted in collaboration with:
Raul K. Suarez, UCSB
Douglas L. Altshuler, UCR
It has been known for some time that the relationship between rates of oxygen
consumption, and ATP production in isolated mitochondria and isolated cells
vary depending upon which metabolic fuel is being oxidized. The latest estimates
of the P/O ratio, the ratio of units of ATP produced to units of molecular
oxygen consumed, indicate that for a given amount of ATP production, fatty
acid oxidation results in approximately 15-18% more oxygen consumption compared
to carbohydrate oxidation.
Despite empirical demonstration of this phenomenon at the level of isolated
cells or organelles and a few examples at the level of isolated organs, it
observed effect on whole animal oxygen consumption rate has never been specifically
recorded. This is a difficult phenomenon to observe in most animals for a
number of reasons. For one, many animals, humans included, do not predictably
and completely shift between fatty acid oxidation and carbohydrate oxidation.
Further, different tissues may be oxidizing differing mixtures of fatty acids
and carbohydrates, and it is difficult to pinpoint the contribution of any
one tissue to overall oxygen consumption rates.
Hummingbirds, however, offer a uniquely well-suited study organism for addressing
the relevance of this phenomenon to whole-animal oxygen consumption. As stated
elsewhere on the website, hummingbirds predictably shift from predominantly
oxidizing fatty acids when fasted, to oxidizing carbohydrates exclusively
during steady-state foraging. In addition, as more than 90% of whole-animal
oxygen consumption during hovering flight is accounted for by the oxygen consumption
of a homogenous population of muscle fibers in the pectoral flight muscles,
extrapolation of the metabolic activity of this tissue type to whole animal
metabolism is comparatively simple. Further, the mechanical cost of hovering
(and thus the required metabolic cost - ATP production) should be unaffected
by the metabolic fuel in use, all else equal.
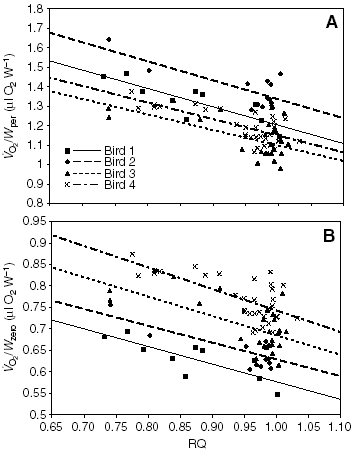
As a result, I hypothesized that, as hummingbirds transitioned from a fasted
(RQ ~ 0.71; oxidizing fatty acids) to a fed (RQ ~ 1.0; oxidizing carbohydrates)
state, the oxygen consumption rate necessary to hold a given mass of hummingbird
aloft would decrease, roughly by 15%. As hummingbirds gained mass while feeding
(transitioning from the fasted to fed state), the cost of hovering would not
strictly remain constant. Thus, I corrected for this changing power output
requirements, and reported the rate of oxygen consumption per unit mechanical
power output. As expected, we discovered that the difference in the rate of
oxygen consumption per unit power output during hovering was between 15 to
18%. This marks the first reported empirical observation of the effects of
P/O ratio on whole-animal oxygen consumption rate.
 Oxygen
consumption rate per unit power output
Oxygen
consumption rate per unit power output