 Hovering Nectarivores: the rufous hummingbird
Hovering Nectarivores: the rufous hummingbird
(Selasphorus
rufus, top), a Panamanian Orchid Bee
(Eulaema meriana, left),
and Pallas' long-tongued
nectar bat (Glossphaga soricina, right).
Fuel Use During Hovering Flight
Research conducted in collaboration with:
Bradley H. Bakken and Carlos Martinez del Rio, U Wyoming
L. Gerardo Herrera M., UNAM (Mexico)
Hovering flight has evolved in only three groups of animals: insects, birds
(hummingbirds), and mammals (glossophagine bats). Within each of these groups,
those species that possess the ability to hover possess a number of traits
in common. All hovering animals are relatively small. Hovering bats and hummingbirds
are among the smallest endothermic vertebrates. Hovering flight is characterized
by high wing beat frequencies and is enabled by some of the highest mass-
specific metabolic rates observed. Notably, all hummingbirds and those
bats that possess the ability to hover are pollinators and largely
nectarivorous.
Is it simply coincidence that virtually all hovering vertebrates are
nectarivorous?
Or, is there a connection between the high metabolic
rates displayed during
hovering and the nature of these animals’ diet?
To investigate fuel use during hovering flight, I have employed a
combination
of respirometric and stable isotope-tracking techniques.
By tracking relative
rates of oxygen (O2) consumption and carbon
dioxide (CO2) production, it is
possible to determine what metabolic
substrates (fat, carbohydrates, or protein)
are being oxidized to support
an animal’s metabolism. A ratio of CO2
produced to O2 consumed (the
Respiratory Quotient, RQ) equal to 0.71 is indicative
of fatty acid
oxidation, while an RQ equal to 1.0 is indicative of carbon dioxide production.
Because, in the case of most exercising vertebrates, the oxidation of protein
makes only a negligible contribution to the support of metabolism, it is possible
to calculate the relative proportion of metabolism supported by either fat
or carbohydrates from the animal’s RQ. RQ values displayed by fasted,
hovering
Hovering Nectarivores: the rufous hummingbird
(Selasphorus
rufus, top), a Panamanian Orchid Bee
(Eulaema meriana, left),
and Pallas' long-tongued
nectar bat (Glossphaga soricina, right).
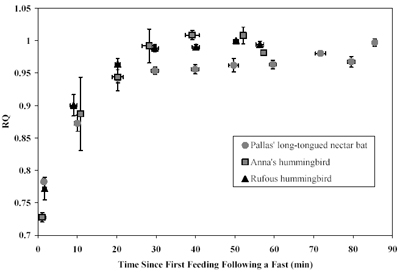 Respiratory
quotient (RQ) during hovering in relation to time
Respiratory
quotient (RQ) during hovering in relation to time
since the first feeding following a fast in Pallas' long-tongued
nectar bats (Glossophaga soricina), Anna's hummingbirds
(Calypte anna), and rufous hummingbirds (Selasphorus
rufus). Animals were allowed to hover-feed at their
own discretion.
bats and hummingbirds are typically low (~0.71-0.78) indicating that fasted metabolism is largely supported by the oxidation of (stored) fatty acids. However, once allowed to feed freely, RQ values from both bats and hummingbirds rise and approach (or meet) 1.0, indicating a switch to nearly exclusive reliance on carbohydrates to fuel hovering metabolism.
RQ values do not allow determination of the source of oxidized carbohydrates or fat. By feeding bats or hummingbirds sucrose solutions with a distinct 13C stable isotopic signature (occurring as a result of differences in the photosynthetic pathway – C3 versus C4 – of the plant from which the sucrose is derived) it is possible to determine the contribution sugar from this exogenous diet towards the fueling of hovering metabolism. This is possible because the 13C stable isotopic signature of expired CO2 reflects the signature of oxidized carbon from which the CO2 derives. This technique, used in combination with measurement of RQ values, has revealed that hovering hummingbirds can support essentially all (average ~ 95%) of hovering metabolism with sugar ingested
no more than 30 minutes prior. Nectarivorous glossophagine bats can support most of their hovering metabolism with recently ingested sugar (average ~ 75%).
The capacity to fuel most or all of hovering metabolism with exogenous sugar
is, based on available studies, an ability restricted to hovering animals.
Humans, by comparison, can support only as much as 30-35% of their exercise
metabolism with dietary sugars. While this is a concrete example of convergent
evolution in the physiology of hovering nectarivores, it remains unclear if
such capacities are displayed solely by these animals or are more generally
associated with small size, high metabolic rates, nectarivory, or the capacity
for hovering flight. More studies are clearly warranted.
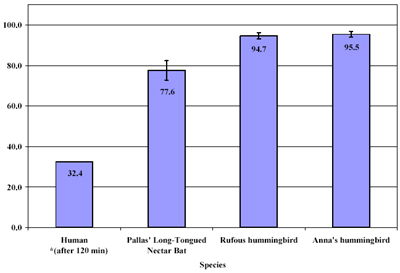 The
proportion of hovering/exercise metabolism supported
The
proportion of hovering/exercise metabolism supported
by exogenous sugars after
30 (120) minutes of access to
sugar for Pallas' long-tongued nectar bat (G.
soricina),
rufous (S. rufus) and Anna's hummingbirds (C.
anna),
and humans. Data for humans from Jentjens et al. (2004)
J. Appl.
Phys. 96:1277.