Evolution of Endomesoderm Specification:
An example of Developmental System Drift
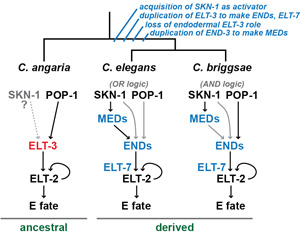
The gut specification pathway in C. elegans and its close relatives involves a cascade of GATA type transcription factors (shown in blue at right). These factors are missing outside of the Elegans supergroup of the genus. We have recently found that gut specification in more distant species occurs through a single ancestral GATA factor, ELT-3 (red). Although ELT-3 is still present in C. elegans, it no longer specifies the gut precursor and instead has roles in the hypodermis and in stress response pathways. The MED, END and ELT-7 factors are very similar in structure to ELT-3, consistent with their having evolved from an ancestral elt-3-like gene by duplication and divergence.
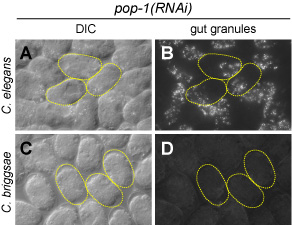
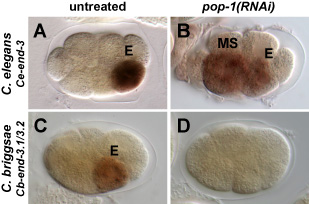
In C. elegans, loss of maternal function of the gene Ce-pop-1 results in a transformation of the MS cell into an E-like cell, causing developmental arrest with an excess of endoderm (gut; Figs. A,B at right). We have found that in the related nematode C. briggsae, depletion of the pop-1 ortholog causes loss of gut (Figs. C,D). This surprising result is actually the 'reverse' of the C. elegans phenotype: Laser ablation studies, in which all cells except E are killed, suggest that E adopts the fate of MS in C. briggsae pop-1(RNAi) embryos.
We
have found that the basis for this difference in pop-1 phenotypes correlates with the way that POP-1 contributes to proper end gene activation in the two species. In both, the end genes are activated in E. In C. elegans, loss of POP-1 results in de-repression of the end genes in the MS cell (and a decrease in their activation overall; Figs A,B on the left), while in C. briggsae, there is complete loss of end expression with Cb-pop-1(RNAi) (Figs C,D).
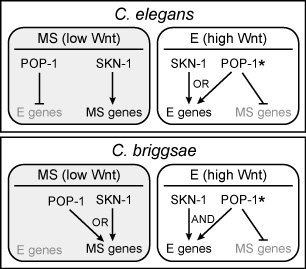
We have also observed that Cb-skn-1(RNAi) results in persistence of MS-derived pharynx, while loss of both Cb-pop-1 and Cb-skn-1 through RNAi results in loss of MS fate. These results show that the regulatory logic of the combined input of the SKN-1 and POP-1 pathways is different in the two species: In C. elegans, SKN-1 specifies MS and E fates, with POP-1 repressing E fate in MS, and contributing to activation of E fate in E; In C. briggsae, SKN-1 and POP-1 provide an 'OR' input into MS fate (with no evidence that POP-1 represses E fate in MS), and an 'AND' input for E fate. Hence, regulation by these factors can be integrated in more than one way to produce the same output.
Other current projects are focused on understanding how the gene network for endomesoderm specification might have evolved and why it underwent expansion, and on how cell fate specification in general has changed within Caenorhabditis and other nematode groups. We have also published a paper that predicts the orthologs of the MED and END factors in the closest relatives of C. elegans: